|
Fósiles del Triasico de
Argentina:
El
Periodo/Epoca triasico abarca un lapso de tiempo desde los 245
hasta 208 millones de ańos antes del presente.
|
 |
Durante
este periodo predomino la tierra firme en vez del mar como
anteriormente y posteriormente, lo que permitió que las
especies animales y vegetales se dispersen por todas las
superficies terrestres del planeta, recordando la
existencia del único supercontinente de Pangea. Las
principales manifestaciones de vida del Triásico en América
del sur proceden principalmente de un sitio fuera de lo
común, un desierto semiárido de la región de Ischigualasto -
Talampaya, en los límites de las provincias de San Juan y
de La Rioja, muy cerca de las cordilleras de Los Andes, en
la llamada vulgarmente como la gran cuenca Triásica de
Ischigualasto - Villa Unión. El área comprende de atractivos
afloramientos de color rojizo, verde y amarillo, de muy
difícil acceso. Estas contienen tres faunas muy diferentes.
La más antigua, de edad Triásico medio, es fauna local de
Los Chańares, donde se hallaron pre-dinosaurios. A estos
niveles se superponen 500 metros de lutitas carbonosas y
areniscas finas de la formación Los Rastros, y sobre ella
continua la formación Ischigualasto, donde se hallaron los
restos de los primeros Saurisquios y Ornitisquios. En el
tope de esta, se halla la formación Los Colorados,
conteniendo fósiles de la última etapa del Triásico
superior. Otro yacimiento muy importante se encuentra en el
sur de la Patagonia, en la Provincia de Santa Cruz <<<
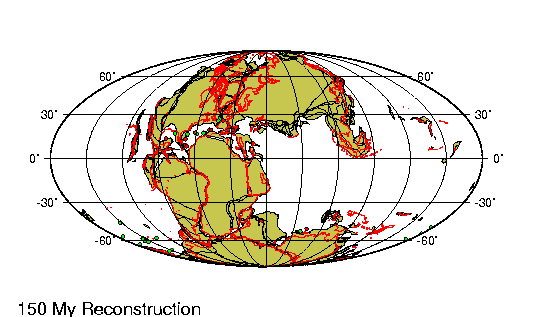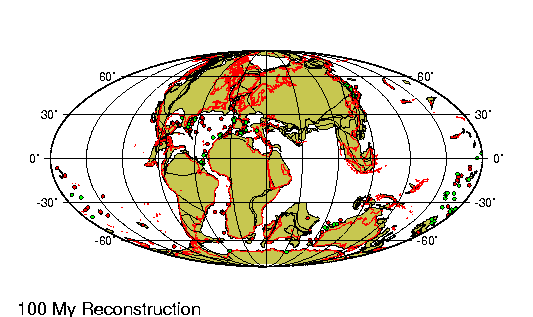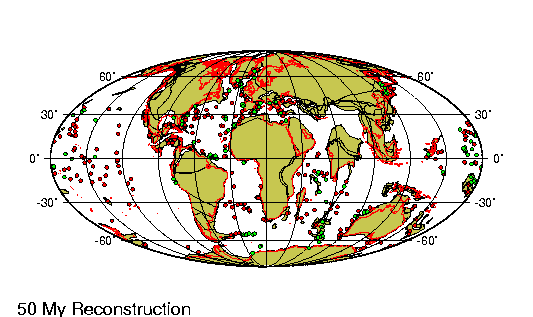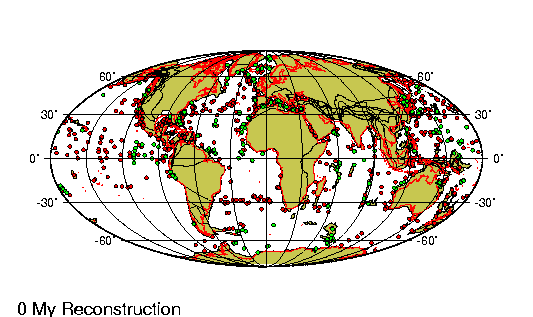
Principales sitios
fosiliferos de Argentina. |
|
Conocida
como estancia El Tranquilito, donde se exponen depósitos
sedimentarios del triasico superior hallándose numerosos
esqueletos de prosauropodos. Los Dinosaurios de Argentina
muestran el verdadero comienzo de la historia evolutiva de
estos grandes animales, aunque para este periodo aun eran
muy pequeńos. Por tal motivo los animales existentes eran
protagonistas de grandes movimientos migratorios, de forma
tal que la fauna mundial era bastante uniforme. Una
situación similar ocurrió con la fauna de la época. El clima
durante este periodo era de carácter subtropical húmedo y la
temperatura global era cálida, aun en altas latitudes. Para
ese momento las capas de hielo se habían derretido ya que
los polos estaban ubicados sobre el océano. A diferencias de
otros periodos geológicos representados en la República
Argentina, no se hallan depósitos sedimentarios de origen
marino. |
Pelorocephalus mendozensis.
Cabrera, 1944.
Anfibio -
tetrápodos. Es un
género extinto de temnospóndilo que vivió a finales del período
Triásico en lo que hoy es Mendoza, Argentina. Presentaba un cráneo
ancho y de contorno parabólico. Su nombre genérico significa "
cabeza monstruosa". Se trata de un género extinto de anfibios
temnosóndilos tetrápodos primitivos, que vivió en el Período
Cretácico Tardío. Su cráneo era triangular - o parabólico -, con
numerosos dientes cónicos y duros, y sus grandes colmillos
palatales, sugieren que se trataba de un depredador. Otras especies
conocidas; Pelorocephalus tenax
(Rusconi, 1949) (originalmente Chigutisaurus
tenax), Pelorocephalus cacheutensis
(Rusconi, 1953) (originalmente
Chigutisaurus cacheutensis) y
Pelorocephalus ischigualastensis?
Bonaparte, 1975.
Taytalura
alcoberi. Martínez, R.N., Simőes,
T.R., Sobral, G. et al. 2021.
|
 |
|
 |
|
Cráneo de uno de los primeros lagartos, el Taytalura
alcoberi, del Triasico de San Juan. |
|
Aspecto de
Taytalura alcoberi, por el paleoartista Jorge Blanco.
|
Lagarto, Lepidosaurio. Aunque otros lepidosaurios
(reptiles con escamas) tempranos se han hallado en rocas de edad
semejante en Europa, Taytalura no solo parece ser
ancestral a todos ellos en su aspecto, sino que es el primero
hallado en el sur de lo que fue el supercontinente Pangea, mostrando
que la evolución del grupo no sólo ocurrió en el norte.. Uno de los
aspectos más importantes del descubrimiento de Taytalura
alcoberi es que permite echar luz sobre el origen del vasto
grupo de especies que incluye a lagartos, lagartijas, serpientes,
anfisbenas y esfenodontes, distribuido por todo el mundo y presente
desde los tiempos de los dinosaurios.. Aunque sólo se haya
encontrado su cabeza, de acuerdo al cálculo de los especialistas, el
animal hallado debe haber medido en total unos quince centímetros de
largo, no muy distinto a cualquier lagartija viviente. Poseía
grandes órbitas y un cráneo ampliado hacia atrás como el de un
camaleón. Sus dientes se hallaban metidos en un largo canal, algo
nunca visto en otros reptiles. Las características anatómicas de
este pequeńo reptil muestran que es un representante temprano de los
lepidosauromorfos, grupo que incluye a los actuales lepidosaurios
(lagartos, serpientes y esfenodontes), que hoy ronda las 10 mil
especies y que tuvo su origen en el mismo momento que los
dinosaurios, los cocodrilos, los pterosaurios y los mamíferos. La
edad del fósil se determinó en unos 230 millones de ańos de
antigüedad, principios del Triásico Superior, una época en la que
los mayores predadores eran parientes terrestres de los cocodrilos y
los primeros dinosaurios iban ganando terreno.
Promastodonsaurus bellmani.
Bonaparte, 1963.
 |
 |
 |
|
Cráneo y mandíbula. (*) |
Cráneo.(*) |
Recreación de Promastodonsaurus.
(*) |
Anfibio -
tetrápodos. Los
Mastodonsauridae es una familia extinta de temnospóndilos,
comprendida entre los capitosauroides. Los fósiles pertenecientes a
esta familia han sido hallados en América del Norte, Groenlandia,
Europa, Asia y Australia. La familia Capitosauridae (también
conocida como Cyclotosauridae y Stenotosauridae) es un sinónimo de
Mastodonsauridae. Su cuerpo tenía el largo de un cocodrilo adulto
actual. Tenía una cabeza chata, con la superficie dorsal ornamentada
con protuberancias, y de un tamańo proporcionalmente grande respecto
al cuerpo.
Pseudochampsa ischigualastensis.
J. Trotteyn y M.Ezcurra, 2012.
 |
|
 |
| Esqueleto de
Pseudochampsa ischigualastensis,
del Triasico del Valle de la Luna. J. Trotteyn y M.Ezcurra,
2012. |
|
Aspecto de
Pseudochampsa ischigualastensis
(*). |
Arcosauromorfo.
Es un género extinguido de archosauriforme, conocido
de la Formación Ischigualasto del Triásico Superior de la provincia
de San Juan, en el noroeste de Argentina. Contiene una única
especie, Pseudochampsa ischigualastensis,
originalmente nombrada como una segunda especie del
Chanaresuchus estrechamente relacionado, basada en un
esqueleto y un cráneo articulados bastante completos. Una revisión
de los restos concluyó que era mejor trasladar especies a su propio
género, ya que no se encontraron rasgos que unieran a P.
ischigualastensis y la especie tipo de Chanaresuchus
con exclusión de otros proterochampsidos. Un análisis
filogenético coloca a ambas especies en una politomía con
Gualosuchus como los miembros más avanzados de
Proterochampsia.
Eran
originalmente similares a los cocodrilos en lo superficial siendo
animales cazadores semiacuáticos de cerca de 1,5 metros de largo,
con una postura de extensión de los codos hacia fuera y hocicos
largos. A diferencia de los robustos contemporáneos terápsidos, los proterosúquidos sobrevivieron
a la catástrofe en el final del Pérmico, quizás porque
eran carrońeros oportunistas, y se refrescaban en el agua durante el
periodo de calor. Cualquiera de estas u otras hipótesis que sea la
correcta, lo que está claro es que estos animales estaban altamente
adaptados en su nuevo ambiente, y se desarrollaron rápidamente. En
el plazo de algunos millones de ańos en el comienzo del Triásico,
los proterosúquidos habían dado lugar al Erythrosuchidae (los
primeros saurópsidos en dominar totalmente su ambiente), que
remplazaron rápidamente a sus antepasados pequeńos y ágiles.
Proterochampsa barrionuevoi.
Reig, 1959.
|
 |
|
 |
|
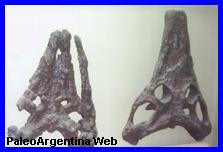
Cráneos de
Proterochampsa. El
primero completo. El siguiente envuelto en sales de hierro.
Museo de La Plata. |
|
Paleoreconstruccion de
Proterochampsa
por Masato Hattori. |
Arcosauromorfo. Es un género de proterosúquidos
encontrado en la Formación de Ischigualasto (Valle
de la Luna) en la provincia de San Juan, es comparable a los
cocodrilos actuales, pero sensiblemente más primitivo en su
estructura anatómica. Pertenece a la familia de los
Proterochampsidae del orden de los Thecodontia. Podía matar pequeńos
dinosaurios que se acercaban a los ríos. Fue descubierto por el
profesor Osvaldo A. Reig en 1959, pertenecía al Triásico tardío,
unos 215 millones de ańos. En el lugar, fue hallado un cráneo
incompleto con mandíbula. Eran delgados, de tamańo mediano de cerca
de 1,5 metros de largo, de largo hocico y superficialmente parecidos
a cocodrilos, aunque carezcan de los osteodermos típicos de éstos, y
en sus características esqueléticas sea mucho más primitivo. Su
característica distintiva es un premaxilar curvado hacia abajo
haciendo que el frente de la mandíbula superior, sobresalga de la
inferior. Los miembros son cortos e indican una postura de
extensión, como los lagartos actuales, pero distinta de la mayoría
de los arcosaurios posteriores. Los proterosúquidos representan
quizás el más temprano ejemplo de radiación adaptativa de los
arcosaurios.
Gualosuchus reigi.
Romer, 1971.
|
 |
 |
|
Cráneo y
mandíbula de la Formación Los Chańares. Copia Col. Museo
Municipal Punta Hermengo de Miramar. |
Aspecto que
presentaría Gualosuchus reigi. |
Arcosaurio.
Las características de este genero son muy
parecidas a aquellas citadas para Chanaresuchus bonapartei.
Su cráneo posee un hocico bajo y alargado, compuesto de una buena
cantidad de dientes cónicos y aserrados para capturar presas. Es muy
probable que pasaría un tiempo muy importante dentro de los espejos
de agua, alimentándose de peces como el descrito mas abajo, o de las
tortugas acuáticas. El largo de esta especie esta calculado entre
los 1,5 a 2 metros de largo y unos 45 centímetros de altura.
Chanaresuchus bonapartei.
Romer, 1971.
|
 |
|
 |
|
Cráneo y mandíbula de
Chanaresuchus (*). |
|
|
Arcosaurio. Es un género
extinto perteneciente al grupo Proterochampsia. El tamańo era
pequeńo para un proterocámpsido, con un promedio de poco más de un
metro de largo. Sus fósiles se encuentran en el Triásico Medio y el
Triásico Superior en la provincia de La Rioja, Argentina y en Río
Grande do Sul, Brasil. Chanaresuchus parece ser uno de
los arcosauriformes más comunes en la Formación Chańares debido a la
abundancia de ejemplares que se han referido al género. Gran parte
del material fue encontrado por la expedición La Plata-Harvard
entre 1964 a 1965. El cráneo es bastante amplio en la parte
posterior con un hocico estrecho, que varía en longitud desde
alrededor de 165 milímetros a 260 milímetros en los ejemplares más
grandes. A diferencia de otros principios arcosaurios
proterocámpsidos, Chanaresuchus tenía poca armadura.
Los osteodermos encontrados son de tamańo reducido, formando una
sola línea en la columna vertebral. Van desde el cuello hasta la
última vértebra presacral y probablemente continuaba por la cola,
aunque no se encontraron especímenes que muestren o sugieran esto.
Hay alrededor de tres escamas por vértebra.
Neoaetosauroides
engaeus.
Bonaparte, 1969.
|
 |
|
 |
|
Esqueleto de
Neoaetosauroides engaeus, en la vieja exhibición del
Museo de Ciencias Naturales Miguel Lilo, Tucumán.
|
|
Posible
aspecto de
Neoaetosauroides engaeus, (*). |
Aetosaurio.
Es un
género extinto de aetosaurio primitivo. Su tipo y la única especie
es N. engaeus. Los fósiles fueron hallados en la formación Los
Colorados que se encuentra a lo largo del cauce del río Sierra
Morada en La Rioja, Argentina, y se remontan a fines del Triásico.
Fue el primer aetosaurio que se descubrió en la formación, los
restos fueron hallados en la década de 1960. El
Neoaetosauroides es uno de los géneros de aetosaurios que se
encuentran mejor representados en Sudamérica, y algunos especímenes
están completos. Los espécimenes encontrados presentan dos filas de
osteodermos dorsales dispuestas a cada lado de la columna vertebral;
además se observan osteodermos ventrales. A diferencia de la mayoría
de los otros aetosaurios, en el Neoaetosauroides el
cuarto dedo es más largo que el segundo y el tercero.
Adicionalmente, el número de falanges en el quinto dígito es mayor
que en otros aetosaurios. La fila superior de dientes del
Neoaetosauroides se dispone por delante del extremo de su
hocico alargado, evidencia que es incompatible con la teoría de un
pico keratinoso propuesta para otros aetosaurios.
Chirotherium higuerensis.
Rusconi 1952.
|
 |
|
 |
|
Icnita o
huella de
Chirotherium, procedente de la Formación
Talampaya, La Rioja. |
|
Aspecto de
Chirotherium higuerensis (*). |
Arcosaurio.
También conocido como Cheirotherium
('bestia de mano'), es el nombre de un rastro o icnofósil triásico
que consta de huellas de cinco dedos (pentadactilo) y pistas
enteras. Estos parecen, por coincidencia, notablemente como las
manos de simios, humanos y osos, con el dedo más exterior
desarrollado para extenderse hacia un lado como un pulgar, aunque
probablemente solo funcione para proporcionar un agarre más firme en
el barro. Las huellas de Chirotherium se encontraron
por primera vez en 1834 en la arenisca del Triásico Inferior en
Turingia, Alemania, que data de hace unos 243 millones de ańos.
Estas huellas fósiles se han encontrado posteriormente en América
del Norte, Argentina, África del Norte, Europa y China.
Las criaturas que hicieron las huellas y huellas eran
probablemente archosaurios pseudosuquios relacionados con los
antepasados de los cocodrilos. Es probable que pertenecieran a
grandes carnívoros con movimientos semi-erectos.
En Argentina se han recuperado algunas huellas de Chirotherium
en el Triásico de la Formación Talamaya (La Rioja) y su localidad
tipo es Las Higueras, que se encuentra en una piedra arenisca /
limolita fluvial, triásica en la Formación Sierra de las Higueras,
Mendoza.
Tarjadia ruthae.
Arcucci et
al. 1998 .
|
 |
|
 |
|
Esqueleto
de Tarjadia en el Museo de Ciencias Naturales de
Ischigualasto. |
|
Aspecto en
vida de Tarjadia ruthae (*). |
Arcosaurio.
Es
una especie extinta
de pseudosuquios
erpetosúquidos , emparentado lejanamente con los cocodrilos modernos
.Es un reptil que vivió en el actual territorio de La
Rioja durante el período Triásico. Los primeros fragmentos fueron
hallados en 1998 y, en la última década, un equipo de investigación
del CONICET encontró cráneos completos y otros restos mejor
preservados. Así, pudo determinar de qué especie se trata y comenzar
a estudiar cuáles eran sus hábitos. Apenas algunos fragmentos de
osteodermos, es decir placas de huesos que cubren la piel, vértebras
y costillas hallados en 1998 en la Formación Chańares, ubicada en el
Parque Nacional Talampaya, hace 237 millones de ańos durante el
período Triásico, le permitieron a las científicas argentinas Andrea
Arcucci y Claudia Marsicano determinar la existencia de una especie
nunca antes descripta. Ese escaso registro fósil no alcanzaba para
establecer con exactitud las relaciones de parentesco de esta
especie y, en ese entonces, las expertas a cargo del hallazgo
consideraron que se trataba de un tipo de reptil muy antiguo
vinculado al origen mismo de los arcosaurios, el linaje que engloba
a los cocodrilos, las aves modernas y muchas de sus formas
precursoras. Debió haber sido un cazador de poca monta. Esto surge
del estudio del oído, donde encontramos que los canales que lo
conforman tienen curvaturas bastante bajas. Esas estructuras son las
que permiten estabilizar rápido la vista y recuperar el balance sin
marearse al percibir movimiento. En este caso, no era así, su
estructura era más primitiva. Entonces no era un animal que
perseguía a sus presas con movimientos ágiles, sino que su forma de
conseguir alimento se basaba en otras estrategias. Tenia entre 2 y 3
metros de largo, T. ruthae era un cuadrúpedo cubierto por osteodermos en el dorso y los miembros, y presentaba un cráneo
robusto en cuyo techo se destacaba una marcada ornamentación,
similar a la que presentan los cocodrilos modernos.
Gracilisuchus stipanicicorum.
Romer, 1972.
|
|
|
|
 |
 |
 |
|
Laja con el esqueleto del
primitivo Guacilisuchus. Colección MMS. |
El técnico Mariano Magnussen Saffer con Guacilisuchus.
|
Aspecto en vida
de Gracilisuchus stipanicicorum del Triasico de Argentina. |
Tecodonte.
De esta especie se conocen varios
ejemplares relativamente completos, caracterizados por presentar un
un tarso cocodriloide (de cocodrilo). Su tamańo era de pequeńas
dimensiones, ya que media 70 centímetros de largo por 30 centímetros
de largo. El cráneo de este pequeńo reptil Arcosaurio es
proporcionalmente bajo y alargado. Su vista dorsal es triangular,
con el extremo anterior muy angosto y la región posterior ancha. Las
grandes orbitas tienen una amplia expresión dorsal, por lo que los
ojos se proyectarían laterodorsal y anteriormente, tal vez con
considerable superposición de campos visuales. Su mandíbula es algo
mas corta que la longitud del cráneo, llegando solo a la mitad del
pre-maxilar. Poseía dos vértebras sacras, mientras que su larga cola
poseía unas 45 vértebras de tamańo decreciente. La extremidad
posterior del tarso es derivado en relación a tecodontes mas
antiguos como Euparkeria, registrado en el triasico
inferior de África. En Gracilisuchus podemos observar
a simple vista que el miembro anterior es 40% menor que las
extremidades posteriores, lo cual es algo muy frecuente entre los
los Ornitosuquidos. Se trato de un pequeńo depredador con ciertas
habilidades, cuyo alimento estaría constituido principalmente de
insectos y vertebrados pequeńos. Para culminar, podemos decir que
Gracilisuchus es un interesante ejemplo de la
diversidad de Tecodontes que surgieron o se diferenciaron durante el
triasico medio, como producto de una gran radiación adaptativa
operada en este grupo de Arcosaurios, y que culminaria con la
diferenciación de ambos ordenes de Dinosaurios (Saurisquia y
Ornitisquios), de los cocodrilos y eventualmente de los Pterosaurios,
que prosperaron a fines del triasico (?) y principios del Jurasico.
Aetosauroides scagliai.
Casamiquela, 1960.
| |
|
|
 |
 |
|
Cráneo y
mandíbula de Aetosauroides.
Copia Col. Museo Municipal Punta Hermengo de Miramar. |
Aspecto de Aetosauroides
(*). |
Tecodonte acorazado. Los
diversos ejemplares disponibles de Aetosauroides corresponden a
especimenes desde unos 80 centímetros hasta 2,50 metros de largo, en
algunos casos con muy buena información sobre el cráneo y poscraneo,
aunque no totalmente completo. El cráneo de esta especie parece ser
mas primitivo que en los Aetosaurios de Europa y América del Norte,
especialmente por ser mas alargados, bajo y ramas mandibulares
extensas y débiles. Sus dientes también son algo mas primitivos, ya
que son pequeńos y agudos en lugar de gruesos y subcilindricos como
en el caso de Typothorax del Triasico superior de
Estados Unidos. Poseían a lo largo de su columna vertebral una tipo de
coraza, formadas de placas paramedianas rectangulares, dispuestas
transversalmente en el sector dorsal, y una hilera de placas
cuadrangulares en el borde o costado lateral. Las mismas presentan
en sus proporciones según correspondan a la región cervical, dorsal
o caudal, pero poseen una decoración uniforme, constante, de
pequeńas fosetas. Algunos huesos de sus extremidades presentan
características propias de los cocodrilos, demostrando que se podían
mover a cierta velocidad. Esta especie de la fauna de Ischigualasto,
compilo un rol muy importante en su nicho ecológico, ya sea
necrófago u omnívoro - herbívoro, que no poseía ningún otro
Tetrápodo de esa fauna. Otras
especies conocidas: A.
subsulcatus Zacarias, 1982 y
A. inhamandensis
Barberena et al., 1985
Sillosuchus longicervix.
Alcober and Parrish, 1997.
|
 |
 |
|
Esqueleto de
Sillosuchus
exhibido en Japón. |
Aspecto de
Sillosuchus
(*). |
Precocodrilo.
Su nombre genérico significa
"ladrón del sur". Este Sillosuchus ladrón de huevos
era un proto-cocodrilo que se deslizaba en dos patas. Es una muestra
del poderío de los cocodrilos durante el Triásico que incluso puso
en riesgo la expansión inicial de los dinosaurios. Fue uno de
los poposáuridos más avanzado que se conocen. Estos animales poseían
un esqueleto muy grácil y liviano, condición esta muy rara para un
animal con tarso de cocodrilo. Posiblemente logró superar la
condición cuadrúpeda de los demás precocodrilos. Sillosuchus
seguramente fue lo suficientemente ágil y veloz como para competir
con los dinosaurios herrerasáuridos. No se han encontrado hasta el
momento especímenes completos, conociéndose solamente parte de la
columna vertebral, pelvis y miembro posterior casi completo.
Venaticosuchus rusconii.
Bonaparte, 1971.
|
 |
|
 |
|
Esqueleto de Venaticosuchus. |
|
Aspecto de Venaticosuchus. |
Diápsido. Es un género
extinto de reptiles saurópsidos crurotarsianos que vivieron a
finales del período Triásico. Fue un depredador de tamańo mediano,
que alcanzaba hasta 2 metros de longitud. Originalmente se pensó que
era el antepasado de los dinosaurios carnosaurios (un grupo que se
pensó abarcaba a todos los terópodos grandes, como
Giganotosaurus); sin embargo, ahora se sabe que están más
estrechamente relacionados con los cocodrilos que con los
dinosaurios. Solo se ha descrito a una especie de
Venaticosuchus, la especie tipo, V. rusconii
del Triásico Superior de Argentina, que data de alrededor de hace
210 millones de ańos. Venaticosuchus es miembro de
Ornithosuchidae, una familia de crurotarsos carnívoros
facultativamente bípedos que estaban geográficamente extendidos
durante el Triásico Superior. Se conocen además a otros dos géneros,
Ornithosuchus y Riojasuchus.
Ischigualastia jenseni.
COX, 1962.
|
|
|
|
|

|
 |

|
|
Esqueleto montado
en el Museo de la Fundación Miguel Llilo de
Tucumán. |
Cráneo de Ischigualastia Jensini.
|
Reconstrucción
en vivo de Ischigualastia Jensini. |
|
Dicynodonte.
Posee un cráneo voluminoso, con la mandíbula gruesa, masiva, que
sugiere una fuerte capacidad para cortar y triturar vegetales. Sus
ramas mandibulares se encontraban recubierta por unos fuertes
músculos. La parte mas anterior del hocico era
proporcionalmente angosta, lo que sugiere que seleccionaría de
alguna manera los vegetales de que se alimentaba. El esqueleto
recuperado y montado en el Museo Pedagógico de la Fundación
"Instituto Miguel Lillo" en la ciudad de Tucumán, nos muestra que su
estructura esqueletaría era muy fuerte, gruesa, conformando un
animal de robustez comparable o posiblemente mayor a la de un
rinoceronte africano actual. Se destacan las cortas y gruesas
extremidades anteriores, con el humero en una posición casi
horizontal y dirigido hacia afuera, lo que posibilito a este animal
apoyar su cuerpo sobre cinco dedos muy fuertes. Las extremidades
posteriores eran totalmente lo contrario, tomando una posición
totalmente vertical, lo que hace pensar a los Paleontólogos que
habría sido mas esbelta en la región posterior y mas voluminosa y
pesada en su mitad anterior, con el cuello corto, grueso y una
pesada cabeza en posición baja, tal vez con algún parecido general a
los búfalos actuales. Procede de los niveles inferiores de la
formación Ischigualasto, ubicado en el "Valle de la Luna", Provincia
de San Juan, Republica Argentina.
Kannemeyeria
argentinensis.
Bonaparte,
1966.
|
 |
 |
 |
|
Cráneo de
Kannemeyeria
argentiensis
(*). |
Cráneo de
Kannemeyeria
(*). |
Aspecto de
Kannemeyeria
argentiensis
(*). |
Dicynodonte.
Es
un género extinto de terápsidos dicinodontos que existió hace 230
millones de ańos en el periodo Triásico. Este herbívoro estaba bien
adaptado a la vida terrestre. Alcanzaba los tres metros de longitud.
Era un herbívoro grande con un poderoso pico y fuertes músculos
mandibulares diseńados para cortar los vegetales que comía. Aunque
tenía una cabeza grande, el cráneo era liviano debido al gran tamańo
de las órbitas y la cavidad nasal. Sus cinturas óseas poseían
grandes placas óseas útiles para soportar su gran corpulencia. Debió
de habitar un ambiente de parajes abiertos, formando manadas, donde
era quizá cazado por depredadores como Cynognathus.
Como sus restos fósiles han sido hallados en Sudáfrica, Argentina,
India, China y Eritrea , podemos suponer que en su tiempo
Kannemeyeria tenía una distribución mundial.
Exaeretodon frenguellii.
Cabrera, 1943.
| |
|
|
|
|
 |

|
|
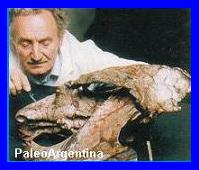
El técnico Sr
Vince, junto a cráneo y mandíbula de Exaeretodon frenguelli. |
Cráneo y
mandíbula de Exaeretodon frenguelli en el Museo de La
Plata. |
Aspecto en
vida de Exaeretodon frenguelli. (*) |
Cinodonte -
Mamiferoide. De Exaeretodon se conocen numerosos restos
procedentes de la formación Ischigualasto, y pertenecen por lo menos
a tres o cuatro especies distintas con características morfológicas
ampliamente diferenciales en las mandíbulas. Los cráneos
coleccionados de este genero oscilan entre 18 y 60 centímetros de
largo total. Observando el cráneo podemos decir que es bastante mas
largo que ancho, con un hocico largo. Posee tres incisivos en cada
pre-maxilar, un canino relativamente grande y una hilera de
pos-caninos. Entre los restos mas importantes rescatados, podemos
mencionar un esqueleto completo de la sección media de la formación
Ischigualasto, el cual tiene unos 2 metros de largo (incluido el
graneo y la cola que son muy cortas). Poseía un tórax voluminoso y
huesos de la cintura escapular y pélvica muy modestos. Estos datos
sugieren que era un animal muy robusto, de patas cortas y gruesas, y
de unos 60 centímetros de alto. Era un animal de movimientos lentos,
aunque si era acechado por algún carnívoro podía ser algo mas ágil
pero por breve tiempo. Es una de las especies mas representativas y
abundantes del Triasico Argentino.
Probelesodon
sanjuanensis.
Martinez & Forster, 1996.
|
 |
|
 |
|
Cráneo
tipo de
Probelesodon sanjuanensis.
(*). |
|
Aspecto de
Probelesodon sanjuanensis. (*). |
Cinodonte -
Mamiferoide. Es
un género extinto de sinápsidos cinodontos de la familia
Chiniquodontidae. Sus fósiles se han hallado en Argentina y Brasil
procedentes del Triásico. Los primeros especímenes se descubrieron
en la formación Chańares en la provincia de La Rioja, Argentina. Una
nueva especie, Probelesodon sanjuanensis, fue descrita
en 1996 basado en la base de un cráneo hallado en la formación
Ischigualasto. La presencia de este género en estos estratos sugiere
que existió durante el depósito de la formación Los Rastros, la cual
posee afloramientos en la misma área donde se han hallado los
fósiles; sin embargo, aún no se han descubierto en esta formación.
Los hallazgos en Ischigualasto están restringidos a los primeros
estratos, extinguiéndose por completo poco después de la
desaparición de los últimos carnívoros cinodontos del área. Esto se
ha tomado como una evidencia del reemplazo gradual de los terapsidos
sustituidos por la fauna dominante de arcosaurios del Triásico
Superior. La evidencia en la formación Chańares también parece
apoyar la hipótesis de la fauna trancisional.
Dinodontosaurus
oliveirai. Romer, 1943.
| |
|
|
 |
 |
 |
|
Coprolito o
excremento fosilizado de Dinodontosaurus. Imagen de
prensa. |
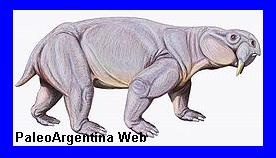
Esqueleto de
Dinodontosaurus. (*). |
Aspecto en vida de
Dinodontosaurus. (*). |
Dicynodonte.
Un
megaherbívoro de ocho metros de largo, similar a los
rinocerontes modernos. Estos animales eran dicinodontes: de
gran tamańo, similares a los mamíferos comunes del período
Triásico, cuando los primeros dinosaurios comenzaron a
surgir.
Los
paleontólogos argentinos han hallado las 'letrinas públicas
más antiguas del mundo', creadas por algunos de los primeros
dinosaurios y que podría servir para esclarecer el misterio
del origen de estas criaturas. Según los científicos, los
miles de excrementos fosilizados desenterrados en la
Formación Chanares, en la provincia argentina de La Rioja,
fueron dejados por grupos de megaherbívoros (mamíferos de
gran tamańo ya extinguidos) similares a los rinocerontes.
el estiércol contiene información muy valiosa para
la paleontología sobre la dieta de
los animales prehistóricos, las enfermedades que padecían
y la vegetación de la época. De acuerdo con el estudio, los
coprolitos (excrementos fosilizados con gran contenido en
minerales inorgánicos) desenterrados son de más de 40
centímetros de ancho y pesan varios kilogramos. Algunos de
ellos son alargados, otros son óvalos de colores desde el
gris claro al marrón y al violeta oscuro. Solo una especie
podría producir bultos de semejante tamańo y también
encontramos sus huesos esparcidos por toda la zona. También
era una advertencia para los depredadores. Los coprolitos no
son nada nuevo, pero fue inesperado y excitante encontrar
una acumulación tan antigua como esta, ya que normalmente
las heces se degradan bastante rápido. Cada excremento es
una instantánea de un antiguo ecosistema, su vegetación y la
cadena alimentaria. Con el nuevo hallazgo los científicos
esperan vislumbrar una imagen del hábitat desaparecido que
dio origen a los dinosaurios.
Gallegosichnus
garridoi. Casamiquela,
1964.
|
 |
|
 |
|
Huellas de Gallegosichnus en una laja de las veredas
de la ciudad de Bariloche. Fuente Ari Iglesias.
|
|
Posible aspecto de
Gallegosichnus (*). |
Reptil Mamiferoide.
En Río Negro son famosas las huellas de unos de los reptiles
más antiguos que se conocen. En la localidad de Los Menucos
(sobre la Línea Sur). De allí son la mayoría de las lajas
que fueron utilizadas para la construcción de la Costanera
de la ciudad de Bariloche. Esas lajas preservan las huellas
de varios reptiles que vivieron en Patagonia hace 200
millones de ańos (Período Triásico). Se trata de pequeńas
huellas de hasta 8 cm de diámetro pero que también preservan
largos caminos de varios metros de largo. Hoy es posible
encontrar muchos de estos fósiles caminando por las veredas
de la ciudad de Bariloche. Algunas huellas preservan la
marca de sus dedos y característicamente se sobre impone el
pie sobre la mano, denotando su neto andar cuadrúpedo (al
retirar la mano, el pie se adelanta y pisa en donde dejó la
huella la mano). Los investigadores reconocieron que estas
huellan deben haber sido producidas por reptiles
plantígrados (que apoyan toda la palma), con importantes
garras tanto en la mano como en el pie. Muy probablemente
correspondieran a un grupo de antiguos reptiles denominados
"reptiles mamiferoides" o reptiles Terápsidos. Otras huellas
conocidas en la localidad de Los Menucos , han sido
adjudicadas a reptiles dicinodontes y a esphenodontes
(tatuaras o camaleones). Otra especie relacionada y bien
registrada es Calibarichnus
Pascualgnathus
polanskii.
Bonaparte, 1966.
|
 |
|
 |
|
Cráneo y mandíbula
de Pascualgnathus (*). |
|
Posible aspecto de
Pascualgnathus (*). |
Cinodonte.
Es un traversodóntido pequeńo. Tenía grandes caninos
superiores y pequeńos dientes postcaninos. Los dientes
postcaninos de Pascualgnathus y otros
traversodóntidos son anchos, permitiéndoles comer material
vegetal. Los postcaninos superiores de Pascualgnathus
son rectangulares. Cada uno tiene un borde central y una
cúspide al lado dirigidos hacia la boca. Hay otras dos
cúspides en un lado del diente dirigidas hacia los labios,
siendo una mayor que la otra. Los postcaninos inferiores
tienen una forma menos rectangular y solo tienen dos
cúspides. A diferencia de los postcaninos superiores, son
más largos que anchos. Se descubrieron restos de
Diademodon en la Formación Río Seco de la Quebrada
junto a Pascualgnathus, lo que sugiere que los
ancestros de Pascualgnathus migraron de África
a América del Sur. vivió durante el Triásico Medio de
Argentina. Sus fósiles se han encontrado en la Formación Río
Seco de la Quebrada.
Chanaria platyceps.
Cox 1968.
|
 |
 |
 |
|
Cráneo, Chanaria
platyceps, en Museo de Ciencias Naturales La
Rioja. |
Coprolitos del Triasico superior de Divisadero
Largo. Museo de Mendoza. |
Aspecto del
sinápsido. (*). |
Dicynodonte.
Es un género extinto de
sinápsidos no mamíferos. Los sinápsidos no mamíferos se han
denominado tradicionalmente "reptiles mamiferoides", es
decir reptiles similares a los mamíferos. El rasgo craneano
que los caracteriza es la presencia de un orificio en los
lados del cráneo tras las órbitas de los ojos, la fosa
temporal inferior. Hace tiempo se supuso erróneamente que
esta fosa se había formado por la fusión de las dos que
existen en los reptiles diápsidos y por ello el grupo
recibió el inadecuado nombre de sinápsidos. Como los
mamíferos, los sinápsidos no mamíferos poseían una piel
glándular carente de escamas, aunque algunos pelicosaurios
conservaron las escamas de los tetrápodos más primitivos en
su parte ventral. Dichas escamas, como el pelo o las plumas,
no tenían la misma estructura que las típicas escamas
reptilianas. Algunas especies sobrevivieron durante el
periodo siguiente, el Triásico, pero no pudieron competir
con los arcosaurios que ocuparon su lugar como vertebrados
dominantes; a finales del Triásico, su número era escaso,
pero para entonces, muchas de las características de la
organización mamaliana se habían desarrollado ya. Durante
los 100 millones de ańos siguiente (los periodos Jurásico y
Cretácico), los sinápsidos vivieron a la sombra de los
grandes saurópsidos (dinosaurios, pterosaurios,
plesiosaurios, etc.) y no alcanzaron tamańos superiores a
los de un gato grande; algunos mamíferos de aspecto
semejante a de los roedores actuales, los multituberculados,
se hicieron abundantes pero se extinguieron durante el
Eoceno.
Massetognathus
pascuali. Romer, 1967
|
 |
 |
 |
|
Restos
de Massetognathus en el Museo de La Plata. |
Cráneo
de Massetognathus (*). |
.Aspecto de Massetognathus (*). |
Cinodonte.
Es un género de cinodontos
traversodontidos, relacionados con Cynognathus.
Fue un herbívoro que vivió en Suramérica durante el periodo
Triásico Medio hace 220 millones de ańos y fue descubierto
en paleorrota (Brasil) y en la formación Los Chańares en
Argentina. Massetognathus tenía cerca de 50 cm
de longitud. Tenía dientes especialmente adaptados para
masticar plantas. Todavía conservaba el hocico largo
distintivo de sus parientes cinodontos, tenía incisivos
cortantes y caninos similares a colmillos, pero los dientes
laterales no eran puntiagudos.
En lugar de ello eran aplanados y estaban
cubiertos por una serie de crestas, lo cual los hacia
adecuados para moler tallos raíces y otros materiales
vegetales. Massetognathus tenía patas con
garras y una cola larga similar a la de un perro. Como la
mayoría de los cinodontos, pudo haber estado cubierto de
pelo.
Pseudotherium argentinus. Wallace.
Martinez y Rowe, 2019.
|
 |
|
 |
|
Cráneo
de Pseudotherium argentinus
(*). |
|
Aspecto de
Pseudotherium argentinus.
Paleoartista Jorge A. González |
Reptil Mamiferoide.
La nueva especie tiene el hocico muy
largo y chato, de poca profundidad, y sus colmillos son
también muy largos y están ubicados casi en la punta del
hocico. En este sitio, también conocido
como Valle de la Luna, del del Triásico esperó a ser
rescatado junto al dinosaurio Panphagia protos, uno de los
sauropodomorfos más antiguos conocidos hasta hoy, y al fémur del
primer y único protodinosaurio lagerpétido descubierto en
San Juan. Medía unos 25 centímetros de longitud
y fue hallada en Ischigualasto
junto a dos de los dinosaurios más antiguos de los que se tienen
conocimiento. Este mamiferoide nombrado como Pseudotherium argentinus
vivió en un ambiente más cálido que el actual, con abundante flora,
compuesto mayormente por helechos y coníferas, ya que todavía no
existían las plantas con flores. Hasta ahora, no se había
encontrado un mamiferoide tan avanzado en
Ischigualasto, en el que se observa el crecimiento de la cavidad
encefálica, precursora de los grandes encéfalos de los mamíferos. Con estas imágenes, se pudo
observar el desarrollado oído interno, la pérdida de la barra posorbitaria, como así también se detectó la presencia de los
turbinales que son como tabiques que le permitieron a este animalito
calentar el aire que ingresaba a su sistema respiratorio, lo
que nos indicaría que tenía sangre caliente.
Scaphonyx sanjuanensis.
Sill, 1970.
|
Reconstrucción, cráneo con mandíbula y esqueleto de
Scaphonyx sanjuanensis, tomados de Bonaparte, 1997. |
Rincosaurio.
Se trata de una especie cuyo largo total oscilaba en los 2 metros
hasta los 2,5 metros de largo, y una aproximada de 45 a 50
centímetros. La posición de las extremidades era primitiva, o sea
con el humero y el fémur próximos a la posición horizontal, lo que
contribuía a la poca altura de los ejemplares. En la formación
Ischigualasto, especialmente en el terciario inferior, la frecuencia
de Scaphonyx
es notable e incluye ejemplares de diversos tamańos que presentan,
muy probablemente, estadios de desarrollo de individuos de la misma
especie. En el Museo de Ciencias Naturales de la Universidad
Nacional de San Juan se encuentra montado un magnifico ejemplar de
esta especie. Es indudable que Scaphonyx sanjuanensis
era una especie totalmente herbívora, y que eventualmente seria una
de las presas de los grandes carnívoros como Saurosuchus
o aun
Herrerasaurus. Los vegetales que probablemente se
alimentara este Rincosaurio es mas difícil estimar. Tal vez la
adaptación de su pico estaba dirigida a arrancar raíces, o
eventualmente para triturar frutos de cicadales según los autores.
Saurosuchus
galilei. Reig, 1959.
|
 |
 |
|
Esqueleto de Saurosuchus
galilei en el parque provincial Ischigualasto |
Recreación en vida de
Saurosuchus galilei. (*). |
Arcosaurio. Es un
género extinto de saurópsido arcosaurio perteneciente al
orden Rauisuchia, perteneciente a la familia
Prestosuchidae. Vivió en el Triásico Superior, en
territorios de la actual Argentina. Con una longitud de
7 metros, seguramente fue el mayor miembro de su grupo,
con la excepción quizás del menos conocido
Fasolasuchus.Al igual que otros rauisuquios,
Saurosuchus caminaba sobre cuatro patas
completamente extendidas. Es probable que fuera capaz de
erguirse sobre las patas traseras por cortos períodos.
Saurosuchus es conocido a partir de varios
esqueletos parciales recuperados de la formación
Ischigualasto de Argentina. El espécimen del holotipo
consiste de un un cráneo completo aunque deformado,
vértebras dorsales, osteodermos dorsales y partes de la
pelvis. Los miembros posteriores, la cola, el cuello y
la escápula se encuentran en varios otros esqueletos. La
especie tipo S. galilei fue nombrada en
1959. Saurosuchus fue también reportado en
la formación Chinle de Arizona en 2002 sobre la base de
dientes aislados y pequeńos fragmentos craneales. El
valor diagnóstico de estos huesos ha sido cuestionado en
estudios posteriores, que los han considerado como de
una especie indeterminada de rauisuquio.
Saurosuchus es uno de los mayores rauisuquios.
El esqueleto completo no es conocido y las estimaciones
de tamańo van desde 6 a los 9 metros de longitud total
corporal.Tenía un cráneo profundo y comprimido
lateralmente. Los dientes son grandes, recurvados y
aserrados. El cráneo es amplio en la parte posterior y
se estrecha en frente de los ojos. El techo craneal y el
maxilar poseen hoyuelos, un rasgo distintivo no visto en
ningún otro rauisuquio.
Riojasuchus
tenuisceps.
Bonaparte, 1969.
|
 |
|
|
|
Cráneo y
mandíbula de Riojasuchus tenuisceps. (*). |
|
Aspecto de
Riojasuchus tenuisceps. (*). |
Arcosaurio.
Es un género
extinto de arcosaurios
cuadrúpedo.
Riojasuchus es miembro de
ornithosuchidae,
una familia de carnívoros
bípedos
facultativos que eran
la extensión geográfica
durante el
Triásico tardío.
Otros dos
géneros, Ornithosuchus
y
Venaticosuchus, se conocen
actualmente.
El espécimen fue encontrado en
la Formación
Los Colorados. El
material consiste en un
cráneo casi
completo y
un postcráneo
parcial. Una
mayoría del material
postcranial
fue preservada. El esqueleto
fue encontrado
con otros en
la sección superior de la
Formación Los Colorados.
Otros que el
holotipo, se conoce
a partir de tres
esqueletos.
El cráneo del
espécimen tipo
es de 25
centímetros de largo,
y tiene una gran
parte
conservado.
La mandíbula
conserva se compone
del maxilar y
la premaxilar.
Vivio
hace
aproximadamente 217
hasta 215
millones de ańos.
Hemiprotosuchus
leali.
Bonaparte, 1969.
|
 |
|
 |
|
Cráneo y
mandíbula de Hemiprotosuchus leali.
Colección del Museo de Municipal Punta Hermengo de Miramar. |
|
Aspecto
semejante. Museo de Comahue. |
Arcosaurio.
Es
un género extinto de
Protosuchidae
de la Formación Los Colorados
de Argentina.
Fue nombrado por el
paleontólogo argentino
José
Bonaparte en 1969.De
la familia de arcosaurios cocodriloformos protosuquios.
Hemiprotosuchus es considerado como el cocodrilo
mas primitivo de América del sur, con una antigüedad de
217 millones de ańos. Poseía una poderosa cola
con la que podía propulsarse mientras se movía en el
agua, aún tenía las patas largas de sus antepasados y
las patas traseras eran mucho más largas que las
delanteras, lo que muestra que surgió de un animal
bípedo. El cráneo es corto y no adaptado totalmente a la
alimentación con peces; el Hemiprotosuchus
probablemente se alimentaba con animales terrestres del
tamańo de una lagartija, su tamańo es de aproximadamente
de un metro de la cabeza a la punta de la cola.
Ejemplares mas modernos al triasico fueron hallados en
Argentina, América del Norte, África y China.
Lagosuchus talampayensis.
Romer,
1971.
|
|
|
|
|

|
 |

|
|
Esqueleto en
laja de Lagosuchus talampayensis en el Museo de
Ciencias Naturales de Miramar. (Reproducción-copia). |
El técnico Mariano Magnussen
Saffer, junto a la laja con el esqueleto de Lagosuchus
talampayensis. |
Reconstrucción
del pequeńo pre-dinosaurio, llamado vulgarmente
como el abuelo de los Dinosaurios. |
Arcosaurio.
Es un género extinto de pequeńos
arcosaurios cuyos fósiles fueron hallados en la Formación Chańares
de Argentina, que data del Triásico Medio. Es considerado por los Paleontólogos como el abuelo de los
Dinosaurios, aunque propiamente no era un Dinosaurio. En realidad se
trata de un genero conocido mas cercano al antecesor hipotetico de
todos los Dinosaurios. Fue un animal bipedo, de pequeńo tamańo,
dotado de un cráneo relativamente reducido, con dientes agudos,
fuertes, propios de un animal depredador. La serie vertebral
presenta ya una zonación característica semejante a la de los
Dinosaurios, con vértebras cervicales proporcionalmente elongadas y
vértebras dorsales con amplias neurapofisis. La morfología del ilion
es algo primitiva, con un proceso preacetabular de escaso
desarrollo. las extremidades posteriores son mucho mas largas que
las anteriores, aproximadamente mas del doble. El pie tiene cuatro
dedos, pero solo apoyaba tres. La estructura general del pie es
grácil, con metatarsales y falanges muy ongadas. Todas las
características mencionadas anteriormente hacen suponer que
Lagosuchus
tenia un marchar de forma digitígrada, es decir, apoyaba solo
los dedos de los pies. Lagosuchus fue un activo
cazador que se alimentaba probablemente de insectos y pequeńos
vertebrados. La gracilidad de su arquitectura esqueletaría indica
que fue un animal agil y corredor. Su habilidad para la marcha se
basa, sobre todo, por presentar un fémur, tibiotarso y metatarsos
largos y notablemente delgados. Además la postura digitígrada
mencionada previamente en el texto, es otra manera de aumentar la
longitud efectiva del miembro posterior para generar una carrera mas
rápida. Algunos Paleontólogos creen que Lagosuchus y
Lagerpeton, podrían ser formas saltadoras. Fue hallado en
capas del Triasico medio de la Formación Los Chańares, en la cuenca
triasica de Ischigualasto - Villa Unión, región limite entre las
Provincias de La Rioja y San Juan, Republica Argentina. Su
antigüedad es de 225 millones de ańos aproximadamente, 6 millones de
ańos antes que los primeros dinosaurios propiamente dichos,
conocidos en la sección inferior del Triasico superior, citados mas
abajo. Genero relacionado: Pseudolagosuchus major.
Lagerpeton chanarensis.
Romer,
1971.
| |
|
|
 |
 |
Arcosaurio.
Fue un pequeńo animal que media
entre 40 y 55 centímetros de longitud. Su dieta seria insectívora y
carrońero ocasional. Una de las características mas llamativas de
Lagerpeton chanarensis, son la longitud de los tres segmentos de
los miembros posteriores, lo que sugiere que era un corredor ágil,
no muy diferente a la especialización de Lagosuchus.
Algunos Paleontólogos sugieren que este
pequeńo animal pudo ser saltador, por estar dotado de un pie
fundamentalmente didáctico, ya que es apoyarían solo dos dedos en el
suelo. La neurapofisis de las ultimas vértebras anteriores de la
región sacra estaban dirigidas hacia adelante, en vez de hacerlo
hacia atrás como en la mayoría de los vertebrados. Esta
característica propia de muchos mamíferos actuales que se mueven a
saltos, pues facilitaba la flexibilidad de la región posterior del
tronco. Esta es una característica interesante en los procesos
evolutivos, cuando nuevos modelos de organización anatómica se
producen y posibilitan una nueva y amplia diversificación de
especies.
Lagosuchus, Lagerpeton y tal vez otra
especie con características morfológicas similares estuvieron
relacionadas, lograron reunir ciertas adaptaciones anatómicas para
dar paso a la evolución explosiva de los Dinosaurios Saurisquios De
este interesante pre- Dinosaurio se conocen una pelvis completa, el
sacro, varias vértebras y sus extremidades posteriores completas,
que proceden de bancos del Triasico medio, en la localidad
fosilífera de los chańares, ya citadas para Lagosuchus,
en el Noroeste de Argentina.
Marasuchus lilloensis.
Sereno y Arcucci, 1994.
Ornithodira. Es un género
extinto de ornitodiros, saurópsidos emparentados con los
dinosaurios, que vivió a mediados del período Triásico, hace unos
230 millones de ańos, en la Formación Chańares situada en La Rioja,
Argentina. Era un pequeńo depredador de solo 40 centímetros de
largo. La especie Marasuchus lilloensis fue descrita
originalmente como una segunda especie de Lagosuchus,
L. lilloensis. Sin embargo, en un estudio de
Lagosuchus realizado por Sereno y Arcucci (1994), los
autores concluyeron que el espécimen original (la especie tipo)
estaba muy mal preservado como para permitir ańadir especímenes
adicionales a ese género. Ellos también notaron que el espécimen de
L. lilloensis tenía proporciones diferentes en las
extremidades con respecto a la especie tipo. Basándose en esto,
ellos asignaron a L. lilloensis a un nuevo género,
Marasuchus.
Pisanosaurus mertii.
Casamiquela, 1967.
Predinosaurio - Silesaúridos.
El único ejemplar conocido hasta hoy, procede de la Hoyada de
Las Lajas, a unos 60 kilómetros al norte de Ischigualasto, en la
Provincia de la Rioja. La pieza tipo presenta un maxilar derecho
incompleto con 12 dientes, una mandíbula con 15 dientes, numerosas
vértebras, cintura escapular y pélvica y algunos huesos largos. El
esqueleto reconstruido mide aproximadamente 1 metro de largo. De
acuerdo a los estudios realizados por la comunidad Paleontológica
Argentina, sabemos que era un animal principalmente bípedo, lo que
es muy notable al observar la diferenciación del largo de sus
extremidades. Los dientes son algo primitivos, seńalando que se
alimentaba principalmente de vegetales blandos, frutos, hojas y
ramas de coristospermas y otras plantas desaparecidas. Los huesos
mandibulares y los circundantes demuestra que Pisanosaurus
mertii
tenia mejillas, lo que le favorecía a la hora de comer, ya que
al masticar las plantas evitaba la perdida de las mismas. Esta misma
característica la podemos observar en otro Dinosaurio Ornitisquio
conocido como Heterodontosaurus de África del sur. Se
pensaba que se trataba del Ornitisquio mas primitivo conocido en todo el mundo.
Hoy sabemos que es un silesaúrido.
Sus
restos se encuentran depositados en el Museo Pedagógico del
Instituto Miguel Lillo de Tucumán. Vivió hace 230 millones de ańos.
Lucianovenator bonoi.
Martínez, R. & Apaldetti, C. 2017.
|
 |
|
 |
|
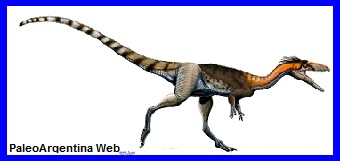
Esqueleto de Lucianovenator bonoi presentado en UNSJ. |
|
Aspecto en vida del terópodo
Lucianovenator bonoi (*). |
Teropodo.
El
hallazgo del nuevo dinosaurio en la Cuenca de Marayes pertenece a
investigadores del Instituto y Museo de Ciencias Naturales de la
UNSJ. Se trata de un espécimen que dio origen a las aves y es el
único en Sudamérica. Su nombre reconoce a un funcionario provincial
y al baqueano cuyo apellido designa el lugar donde fueron
encontrados estos fósiles: Balde de Leyes, o el sitio paleontológico
del nuevo siglo. Si Ischigualasto es el signo de los descubrimientos
paleontológicos en San Juan, el país y el mundo en el Siglo Veinte
-aunque su territorio sigue siendo un riquísimo potencial de
hallazgos-, sin dudas el sitio Balde de Leyes, en el departamento de
Caucete, es el signo del Siglo 21. Porque ni bien comenzado el
milenio actual, los investigadores de la Universidad Nacional de San
Juan empezaron a descubrir allí fósiles de más de cien especímenes,
incluidos reptiles voladores y tortugas, que fueron ampliando la
documentación paleontológica hasta llegar a la actual joya: el
Lucianovenator bonoi. Es un bípedo, cazador y carnívoro,
seguramente un animal muy ágil, de unos dos metros de largo y de un
peso de alrededor de 45 kilos. Existieron durante el Período
Triásico alto, la parte más nueva de ese periodo, antes de las
extinciones de animales entre el Triásico y el Jurásico, suponemos
que tiene entre 205 y 210 millones de ańos. Posee un cuello muy
largo para ser carnívoro. Los dinosaurios carnívoros generalmente
tenían cuellos cortos. Además, esta especie tiene en las vértebras
del cuello cavidades llamadas Pleurocoelus. Esas cavidades
alivianan, pero por otro lado por allí pasan ‘sacos de aire. El
Lucianovenator bonoi pertenece a una familia denominada
Coelophyísido. Es el primer registro de esta familia en Sudamérica.
Se los conocía en África, América del Norte y Asia.
Herrerasaurus
ischigualastensis.
Reig, 1963.
| |
|
|
|

|
 |

|
|
Esqueleto de Herrerasaurus. En el MACN. |
Cráneo y
mandíbula de Herrerasaurus ischigualastensis. (*) |
Reconstrucción
del primitivo dinosaurio terópodo Herrerasaurus
ischigualastensis.(*) |
Teropodo. Un esqueleto muy completo y articulado fue
descubierto por un equipo de Paleontólogos de la Universidad
Nacional de Tucumán, como así también se resguardan otros restos
incompletos de la misma procedencia estratigráfica. A pesar de que
se trata de una forma primitiva de Dinosaurio Saurisquio, el mismo
media unos 3,5 metros de largo. Su majestuoso cráneo mide unos 30
centímetros y presenta un hocico bien desarrollado, que se encuentra
representado mas de la mitad del mismo cráneo. Presenta una larga
hilera de 20 dientes recurvados y puntiagudos que se
complementaban con una garra lacerante en la mano. Dado a los
últimos estudios realizados, imaginamos que Herrerasaurus
fue un saurisquio muy activo, para correr muy hábilmente y depredar
a Rincosaurios y Cynodontes omnívoros. Este Dinosaurio carnívoro
procede de las acumulaciones sedimentarias de Ischigualasto,
Provincia de San Juan, Argentina. Posee una antigüedad de 230
millones de ańos. Arriba: esqueleto montado en el Museo Pedagógico
de San Miguel de Tucumán.
Frenguellisaurus
ischigualastensis.
Novas, 1986.
|
 |
|
 |
|
Museo de la
Universidad Nacional de San Juan. (*) |
|
Posible
aspecto de Frenguellisaurus (*) |
Teropodo.
El dinosaurio depredador más grande conocido en Ischigualasto del
período Triásico Tardío Cárnico, llegando a un largo de 7 metros y
un cráneo de 60 centímetros de largo. Hasta la fecha se ha
descubierto un solo ejemplar, y no se descarta la posibilidad de que
se trate de un Herrerasaurus adulto viejo. Su
procedencia es de la formación Ischigualasto - Valle
de la Luna, San Juan (Argentina). Fue el depredador más temible de
la fauna de ischigualasto. Ágil corredor. La fuerza de sus miembros
anteriores les permitía cazar a sus presas con facilidad. Se conocen
fragmentos de cráneo y mandíbula; parte del miembro anterior y cola
casi completa. Se colectó sólo un ejemplar.
Sanjuansaurus gordilloi.
Alcober y Martínez, 2010.
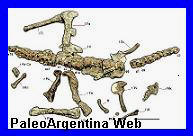
|
 |
 |
 |
|
Ubicación in situ de los
restos encontrados. (*) |
Fragmento de cráneo y
maxilar. (*) |
Posible
aspecto de Sanjuansaurus. (*) |
Teropodo.
Es un género representado por una única especie de dinosaurio
saurisquio herrerasáurido, que vivió a finales del período Triásico,
hace aproximadamente 231 millones de ańos, en el Carniano, en lo que
hoy es Sudamérica. Sus restos se encontraron en la Formación
Ischigualasto de la provincia de San Juan, en el noroeste de
Argentina. Se lo conoce a partir del holotipo, un esqueleto parcial
asociado y parcialmente articulado, PVSJ 605. El mismo consiste en
un fragmento del maxilar, la mayoría de las vértebras desde el axis a
la parte anterior de la cola, la escápula, un cúbito, parte de la
pelvis, la mayoría de los huesos largos de las piernas y algunos
otros huesos . PVSJ 605 fue encontrado en el miembro Cancha de
Bochas cerca de la base de la Formación Ischigualasto, en el Parque
Provincial Ischigualasto. Este horizonte se remonta a
aproximadamente 231,4 millones de ańos atrás, a finales del Carniano
durante el Triásico tardío. Sanjuansaurus fue nombrado
y descrito en 2010 por Oscar Alcober y Ricardo Martínez. La especie
tipo es S. gordilloi en honor a Raúl Gordillo, jefe de
los preparadores de fósiles y artista del laboratorio del Museo de
San Juan. Sanjuansaurus fue contemporáneo de otros
dinosaurios tempranos como Chromogisaurus,
Eoraptor, Herrerasaurus y Panphagia.
Alcober y Martínez realizaron un análisis filogenético en el que
encuentran a Sanjuansaurus dentro de los
herrerasáuridos. Es comparable en tamańo a un Herrerasaurus
mediano, con una longitud del fémur de 395 milímetros, por lo que
probablemente llegó a medir alrededor de 3 metros.
Sanjuansaurus fue descubierto en 1994 por el paleontólogo de
la Universidad Nacional de San Juan Ricardo N. Martínez. En un
principio fue considerado otro ejemplar de Herrerasaurus.
Durante su preparación y limpieza en el laboratorio del Museo de
Ciencias Naturales de San Juan, los investigadores se dieron cuenta
que se trataba de un nuevo tipo de dinosaurio basal.
Eoraptor lunensis.
Sereno et al 1993.
 |

|
 |
|
Aspecto del cráneo del
pequeńo dinosaurio Eoraptor lunensis. (*). |
Esqueleto de Eoraptor.
Museo Argentino de Ciencias Naturales de Buenos Aires. |
Reconstrucción de su posible
aspecto en vida de Eoraptor lunensis. (*). |
|
 |
|
Esqueleto y reconstrucción de
Eoraptor lunensis. Museo
de Ciencias Naturales y Antropologicas de Mendoza. |
Teropodo.
Eoraptor se aproxima, por estructura y por tamańo, al
supuesto antecesor común de todos los dinosaurios. Además, su cráneo
no tiene ninguna de las especializaciones que caracterizan a los
tres grandes grupos de Dinosauria (Ornithischia, Sauropodomorpha y
Theropoda). Presenta como carácter único una dentición heterodonta:
los dientes anteriores de la mandíbula superior (dientes
premaxilares y maxilares anteriores) tienen una forma
característica, expandida, similar a la de los dinosaurios
herbívoros y diferente del resto de los dientes, que son recurvados
y serrados, típicamente teropodianos. Sin embargo, exhibe una serie
de rasgos que lo identifican como el terópodo más primitivo
conocido. Por ejemplo, aunque presenta cinco dedos en la mano, los
dos más externos están tan reducidos que sólo los otros tres serían
funcionales. Estos tres dedos tienen una serie de caracteres
destinados a aumentar la capacidad de la mano para apresar y
manipular el alimento: el dedo II es el más largo, las garras son
curvadas, tiene capacidad de hiperextensión de los dedos, etc. La
mano tiene un gran tamańo respecto al resto del brazo. Además, los
huesos largos son huecos, como en el resto de los terópodos. Así
pues, podemos considerar a Eoraptor como un animal que
ejemplifica al dinosaurio primitivo tipo en el que aparecen ciertos
rasgos que lo definen como el terópodo más primitivo conocido. Modo
de vida: Eoraptor
habría ocupado el nicho de un pequeńo depredador,
alimentándose de reptiles, insectos, etc. y huiría ante la presencia
de los grandes carnívoros, Herrerasaurus incluido, que
lo considerarían como una presa. Sin embargo, la posición
parasagital de sus miembros posteriores, junto a su condición
bípeda, le convertirían en una presa rápida y ágil, difícil de
atrapar por los grandes depredadores de la época. A su vez, esta
rapidez y su agilidad serían grandes ventajas a la hora de capturar
los esquivos animales que constituirían su alimento.
Zupaysaurus
rougieri.
Arcucci & Coria, 2003.
 |
 |
|
Cráneo del depredador Triasico Zupaysaurus. |
Reconstrucción de
Zupaysaurus
rougieri. |
Teropodo.
Hallado en
1996 en la riojana Quebrada de los Jachaleros. Era
como un diablo de paso veloz, que corría detrás de sus presas por
las quebradas de la actual provincia de La Rioja, Republica
Argentina. Para todo aquel desafortunado que se haya cruzado en su
camino, la extrańa cresta sobre su hocico y el centenar de pequeńos
y filosos dientes que poblaban sus temibles fauces no podían pasar
inadvertidos.
A la hora de ponerle un nombre, los paleontólogos se
inclinaron por Zupaysaurus. Zupay es una palabra de origen
quechua que significa diablo o demonio y que se ajusta muy bien a
este dinosaurio de aspecto realmente feroz. Habitó parte del noroeste argentino durante el Triasico,
hace 220 millones de ańos. Desde un punto de vista evolutivo se lo
considera un carácter primitivo, mientras que el resto del animal
presenta caracteres más derivados o modernos que los del resto de
los dinosaurios carnívoros que vivieron en la misma época. Está a mitad de camino entre los dinosaurios
primitivos como el Herrerasaurus y los más modernos como el
Alosaurus . Justamente, la familia a la que pertenece está
constituida por dinosaurios que se especializaron en la locomoción,
pudiendo desarrollar la velocidad de un caballo.
Con sus rasgos modernos, el aún primitivo Zupaysaurus
rougieri
puede
ser considerado el primer tetanuro, familia que incluyó a
dinosaurios carnívoros típicos del jurásico (período posterior
al triásico superior, al que pertenece esta suerte de diablo) y que
llega hasta nuestros días a través de sus lejanos descendientes:
las aves. Pero que el tataratatarabuelo de todos los tetanuros haya
asomado su hocico en el triásico propone todo un cambio de
pensamiento para la paleontología. Esto,
a su vez, implica tirar hacia atrás también la aparición de otras
formas, como los terópodos o los saurópodos, que podrían haberse
originado antes de lo que se piensa, pues se necesita mucho tiempo
de evolución para pasar de una formas a otras. Por otro lado,
el Zupaysaurus constituye un elemento que permite comenzar a llenar
un vacío en la evolución de los dinosaurios Argentinos. Si bien en
el resto del mundo la fauna del triásico superior está muy bien
representada por los distintos hallazgos realizados, en la región
había un vacío muy grande.
Ischisaurus cattoi.
Reig, 1963.
Teropodo.
Su nombre genérico significa " reptil de Ischigualasto ", por
haber sido hallado en esta ( zona ), y el específico, fue dado en
honor a la dra. Noemi Cattoi, del Museo Argentino de Ciencias
Naturales, por su colaboración. Se trata de un saurisquio terópodo -
carnívoro bípedo - herrerasáurido, que vivió en el Período Triásico
Tardío, hace 210 a 228 millones de ańos, en el Carniano. Medía 2
metros de largo, y apenas sobrepasaba el metro de altura. Tenía
musculosos miembros posteriores, y los anteriores eran cortos. Fue
hallado en Ischigualasto, en 1963, por Osvaldo Reig.
Eodromaeus murphi.
Martínez et al., 2011
Teropodo.
Pequeńo, de
unos 5 kilos de peso y poco más de 1,2 metros de alto, rápido y
carnívoro, el Eodromaeus murphi o "corredor del alba"
vivió hace 230 millones de ańos en Argentina. Los científicos que
acaban de descubrir este dinosaurio creen que podría ser el
antecesor del voraz Tiranosaurio Rex o del Giganotosaurus
carolinii. Un grupo de paleontólogos halló los primeros
restos de esta criatura en 1996, en el Valle de la Luna, en el
nororeste de Argentina, como explican en un artículo publicado en la
revista Science. Por
fin, en el ańo 2011, han conseguido recomponerlos. El animal
pertenece al Triásico tardío y, según explican los investigadores,
es uno de los dinosaurios más antiguos hallado hasta ahora, cuya
evolución en la familia de los terópodos, es probable que culminase
en temibles depredadores. Su nombre: "corredor el alba", hace
referencia a que la aparición de este espécimen se sitúa en "el alba
de la aparición de los dinosaurios". Era un animal de dos patas y
pequeńo. Era ágil, lo sabemos por el pequeńo tamańo de sus
extremidades, y tenía manos con capacidad de agarrar cosas con
garras muy poderosas. Fue un depredador, lo sabemos por sus manos,
pero especialmente por sus dientes curvos y largos. Era un carnívoro
especializado y en muchos aspectos es un animal muy cercano al
primer carnívoro, el primero en la línea de los terópodos que
eventualmente evolucionaría y adquiriría tamańos tan enormes. Los
huesos fosilizados del Eodromaeus murphi
aparecieron en unas rocas próximas a los Andes. La mayoría
estaban recubiertos de incrustaciones de hierro y ha sido necesario
un minucioso trabajo para recomponerlos en moldes y reconstruir el
esqueleto completo.
Riojasaurus incertus. Bonaparte,
1969.
| |
|
|

|
 |
|
El técnico Mariano
Magnussen Saffer, junto al esqueleto de Riojasaurus incertus, exhibido en el Museo
de Ciencias Naturales del Instituto Miguel Lillo, Tucumán
(2012). |
Riojasaurus
incertus, reconstrucción. Tomado de Bonaparte, 1997. (*) |
Prosauropodo. Es un representante primitivo de
Dinosaurios Prosauropodos, frecuentemente registrados en la sección
superior de las capas de Los Colorados. El tamańo de los especimenes
recolectados en distintas campańas Paleontológicas, varían de 2 a 7
metros de largo. Se conoce solo un cráneo bien preservado, el cual
mide solo unos 25 centímetros de largo, es muy bajo y alargado,
angosto en vista dorsal y hocico alargado. Su mandíbula inferior
presenta una hilera de dientes y una probable estructura cornea en
forma de pico. Al igual a otros Prosauropodos y Sauropodos
posteriores, se caracterizaba por poseer una cabeza pequeńa, cuello
y cola muy largos, y totalmente cuadrúpedos. Eran de hábitos
herbívoros, alimentándose principalmente de plantas altas (Dicroidium)
y follaje. Es uno de los dinosaurios mejor conocidos del Triasico, y
algunos de sus parientes mas próximos fueron hallados en zonas tan
distantes como Europa, África y China. Su procedencia es de La
Quebrada de los Jachaleros, Provincia de la Rioja, Republica
Argentina y vivió hace 225 millones de ańos.
Coloradisaurus brevis.
Bonaparte, 1978.
 |
|
 |
| Cráneo y
mandíbula de Coloradisaurus.(*) |
|
Coloradisaurus
en el Sendero del Triásico - Parque Nacional de
Talampaya (La Rioja) |
Prosauropodo. Es
un género representado por una única especie de dinosaurios
sauropodomorfo masospondílido, que vivió a finales del período
Triásico, hace aproximadamente 218 y 210 millones de ańos, en lo que
es hoy Sudamérica. Coloradisaurus fue un prósauropodo
de tamańo medio que llegó a medir 4 metros de largo, 2 de alto y
unos 100 kilogramos. La cabeza es alta y corta con órbitas oculares
grandes. Los orificios nasales están bien desarrollados. La región
de la abertura bajo temporal es similar a la del plateosaurio, pero
de menor tamańo. Los restos pertenecen a la Formación Los Colorados
de la provincia de La Rioja, en la Argentina. La expedición que
realizó el hallazgo fue organizada por la Fundación Miguel Lillo,
con el soporte financiero de Consejo Nacional de Investigaciones
Científicas y Técnicas. Martín Vince, Juan C. Leal, Tomás H. Fasola
y José F. Bonaparte participaron en esta. Existe la posibilidad que
Coloradisaurus sea un ejemplar adulto de
Mussaurus. Los últimos hallazgos sugieren que
Coloradisaurus es el taxón hermano de Lufengosaurus
y ambos pertenecen claramente a Massospondylidae.
Mussaurus patagonicus.
Bonaparte & Vince, 1979.
|
|
|
|
|

|
 |
 |
|
Esqueleto
articulado de un ejemplar juvenil depositado en el Museo
Argentino de Ciencias Naturales de Buenos Aires. (*). |
Esqueleto de
Mussaurus patagonicus comparado con una moneda. (*). |
Reconstrucción del nido del
dinosaurio prosauropodo Mussaurus, hallado en la provincia
de Santa Cruz. Dibujo de Irma Dariozzi. Tomado de Bonaparte
(1984). |
Prosauropodo. Saurisquio primitivo
de hábitos herbívoros, conocidos como Sauropodos,
caracterizados por una talla monedadamente grande, pero sin el
tamańo gigantesco de sus parientes del Jurasico y Cretácico. El
material colectado de Mussaurus corresponde a la
Provincia de Santa Cruz, en capas del Triasico superior. Se
descubrieron ejemplares de distinta talla, desde individuos
aparentemente recién nacidos hasta ejemplares adultos, los cuales,
superaban los cinco metros de largo. Poco después una comisión de
investigadores hallaron en la misma zona un conjunto de ejemplares
muy pequeńos, y asociados a ellos un conjunto de huevos fosilizados,
lo que representa el nido de Dinosaurio mas antiguo del mundo. Los
resultados de los estudios indica que se trataba de un nido
atribuido a esta especie donde "recién" habían nacido entre ocho y
diez crías y dos huevos sin eclosionar. Asimismo posibilito realizar
estudios del desarrollo ontogenetico (crecimiento) de este grupo.
Entre fines del periodo Triasico y principios del Jurasico los
Prosauropodos fueron reemplazados por los Sauropodos los cuales
alcanzaron grandes dimensiones.
Panphagia protos.
Martínez & Alcober, 2009.
Prosauropodo.
Era dinosaurio
omnívoro que vivió hace 228 millones de ańos, considerado el
ancestro más antiguo de los herbívoros gigantes y un eslabón perdido
con los carnívoros y de cuatro patas. Este animal es una pieza muy
importante en el rompecabezas sobre el origen de los dinosaurios. El científico formó
parte de la expedición que hace tres ańos halló los restos en el
Parque Ischigualasto-Valle de la Luna, al noroeste del país, donde
se realizaron las investigaciones para determinar su naturaleza. La información que se tenía, hace suponer
que el antepasado más antiguo de los gigantes herbívoros, cuyos
fósiles se habían encontrado, era el
Satrurnalia
tupiniquim, hallado en
Brasil e investigado en 1999, pero el descubrimiento de
Ischigualasto
echa por tierra lo que se creía. El eslabón perdido
fue bautizado 'Panphagia protos' y se trata del nexo
primitivo entre los bípedos carnívoros y los gigantes herbívoros de
cuatro patas, denominados saurópodos, que vivieron en los períodos
jurásico y cretácico de la era mesozoica. Panphagia
significa en griego 'que come de todo' y protos 'el primero. A diferencia de sus sucesores, el 'Panphagia
protos' tenía 1,5 metros de extensión y 30 centímetros de alto,
y era omnívoro, con una dentadura adaptada para alimentarse tanto de
carne como de vegetales.
Adeopapposaurus
mognai.
Martínez, 2009.
|
 |
 |
Prosauropodo.
żCómo fue que los pequeńos dinosaurios
carnívoros, de a lo sumo un metro y medio de largo, llegaron a
convertirse en animales herbívoros gigantescos, con un peso
equivalente a 20 elefantes? La respuesta, que implica el estudio de
millones de ańos de evolución, acaba de dar con un punto clave. Se
trata del Adeopapposaurus mognai, un dinosaurio de 200
millones de ańos de antigüedad cuyos restos fósiles el equipo de
paleontólogos del Museo de Ciencias Naturales de la UNSJ. El
hallazgo se produjo en la localidad Jachallera de Mogna y lo más
valioso que tiene este animal es su pico, ya que agrega una nueva
hipótesis a las teorías sobre su desarrollo.
Hasta ahora, lo que se manejaba en el campo
científico de todo el mundo era que los sauropodomorfos habían
llegado a ser herbívoros gigantes gracias a tres elementos
evolutivos clave: el desarrollo de piedras en el estómago para
triturar las fibras de los vegetales, los dientes adaptados para
cortar esas fibras y el largo del cuello, que les permitía alcanzar
las plantas más altas. De hecho, Adeopapposaurus
significa "lagarto que come lejos" en latín, por su cuello tan
largo. Y mognai hace referencia al lugar donde se lo
encontró.
Por otro lado, haber encontrado estos
restos tan bien conservados en Mogna (y no en Ischigualasto, cuyo
suelo corresponde a sedimentos del Triásico) también es importante
porque Adeopapposaurus es el primer hallazgo de
dinosaurios del Jurásico Inferior para toda Sudamérica y Mogna es el
primer ejemplo de Jurásico en zona de precordillera. Por eso este
descubrimiento e investigación del museo y de la Facultad de
Ciencias Exactas de la UNSJ es fundamental para el mundo de la
ciencia.
Ingentia prima.
Cecilia Apaldetti et al., 2018.
|
 |
|
 |
|
Tareas de
campo para recuperar lo restos de
Ingentia
prima. Imagen de prensa.
|
|
Posible
aspecto el sauropodo Triasico
Ingentia
prima. Imagen de prensa.
|
Sauropodo.
Es un dinosaurio saurópodo herbívoro de gran
tamańo, la única especie del género extinto Ingentia.
Vivió a finales del período Triásico en el centro-oeste de América
del Sur.
Investigadores argentinos presentaron a Ingentia
prima, el primer dinosaurio gigante que habitó el Planeta hace
más de 200 millones de ańos. Supera tres veces el tamańo de los
dinosaurios más grandes del Triásico conocidos hasta ahora. El
hallazgo se produjo en el yacimiento de
Balde de Leyes, al sureste de la provincia de San
Juan.
Los dinosaurios no siempre fueron gigantes. A la
historia evolutiva le llevó millones de ańos para que algunas
especies duplicaran el peso de un elefante actual y alcanzaran entre
ocho y diez metros de largo, pero ese tiempo fue mucho menor al que
se creía: de allí la gran importancia del hallazgo de Ingentia
prima, que habría tenido una masa corporal de hasta diez
toneladas. Antes de este descubrimiento, se consideraba que el
gigantismo había surgido durante el período Jurásico, hace 180
millones de ańos aproximadamente, pero Ingentia prima
vivió a fines del Triásico, entre los 210 y 205 millones de ańos.
Se encuentra depositado en la colección del Instituto de
Paleovertebrados y Museo de Ciencias Naturales, Universidad Nacional
de San Juan, ciudad de San Juan, Argentina. Se calculó que en vida
los ejemplares de esta especie medían entre 8 y 10 metros de
longitud y pesaban cerca de 10 toneladas.
Palaeochersis talampayensis.
Rougier, de la Fuente & Arcucci, 1995.
|

|
 |
|
Ejemplar de
Palaeochersis talampayensis, preparada por M.
Isasi y estudiada por G. Rougier y M. de la Fuente MACN. |
Aspecto de
Palaeochersis,
la primitiva tortuga de Triasico
de Talampaya, |
Reptil. El registro fósil de
tortugas en América austral es muy rico e incluye localidades en el
sur de la Patagonia donde las condiciones climáticas actuales
impiden el establecimiento de quelonofauna. Pero el registro más
antiguo corresponde a Palaeochersis talampayensis, del
Triásico superior del noroeste argentino, en la actual provincia de
La Rioja. Esta tortuga data entre 210 y 220 millones de ańos.
la
subclase Anapsida, caracterizada por un techo craneano contínuo, no
interrumpido por fenestras en la región temporal. Algunas de sus
formas de tortugas más recientes pueden exhibir sin embargo un
cráneo secundariamente modificado, extendiéndose en cada lado de la
cabeza un hundimiento que desde su parte posterior hacia adelante
separa el hueso escamoso del parietal. Las placas córneas que
involucran el caparazón no coinciden en número, ni en forma, con los
huesos colindantes y tampoco con sus suturas. Son menos numerosas y
se subdividen dorsalmente en placas o escudos centrales, o
vertebrales; placas costales o laterales; placas marginales.
Ventralmente hay pares de placas irregulares y anchas, indicadas
como gulares, humerales, pectorales, abdominales, femorales y
anales.
Palaeochersis talampayensis,
procede de la Formación los colorados, ubicados en el Valle de
Talampaya, en la Provincia de la Rioja, Argentina. Otra tortuga
conocida es
Waluchelys cavitesta.
Yelaphomte praderioi.
Martínez et al. 2022.
|
.jpg) |
|
.jpg) |
|
Fragmento mandibular del
reptil volador del Triasico de San Juan, Argentina. |
|
Aspecto de los reptiles
voladores mas antiguos encontrados. Por el Paleoartista
Jorge Blanco. |
Reptil volador. Luego de medio
siglo de una incertidumbre no resuelta por la ciencia en el mundo
acerca del origen y evolución de los pterosaurios, un hallazgo
sanjuanino acude ahora con las respuestas. Esta especie de
pterosaurio medía no más de 30 centimetros en total y tenía una
enorme cresta que sobresalía sobre el pico. Ricardo Martínez lo
bautizó así por la voz en allentiac (lengua huarpe) equivalente a
"bestia del aire" y por Ángel Praderio, quien descubrió el fósil en
Balde de Leyes durante la campańa realizada por el Museo de Ciencias
Naturales de la Universidad Nacional de San Juan en 2014. Los
pterosaurios fueron los primeros reptiles voladores que habitaron la
Tierra desde hace unos 220 millones de ańos, en el supercontinente
conocido como Pangea, previo a los movimientos que llevaron a la
división geográfica actual de cinco continentes. No derivaban de los
dinosaurios, sino que compartían con ellos un ancestro en común. En
estos sedimentos también se descubrió a Pachagnathus benitoi.
Mucho más grande que el anterior pterosaurio, medía hasta un metro y
medio de largo. Su nombre significa "mandíbula de la tierra"
(también del allentiac) y homenajea además al guía Benito Leyes.
fueron clave para determinar los caracteres únicos de las especies a
las que pertenecieron. Ambos reptiles voladores, convivieron en esa
zona desde hace al menos unos 205 millones de ańos.
Triasseuryptilon
acostai. Marquat, 1991.
|
 |
|
 |
|
Linfa de plecóptero del Triasico del Cerro Cacheuta,
provincia de Mendoza. |
|
Reconstrucción en vida de la ninfa de
plecoptera sobre una hoja de Dicroidium bajo la superficie
del agua (dibujos de Carsten Brauckmann y Elke Gröening,
2009).
|
Insecto, Neoptera. Los plecópteros son insectos poco comunes y se encuentran cerca
de lagos o vertientes. Poseen una
metamorfosis incompleta o
hemimetabolismo donde los estados
ninfales son acuáticos y los adultos voladores. Las ninfas son
aplanadas, con
aparato bucal masticador y tienen largos
cercos y
antenas. Los adultos son relativamente grandes, con cuatro
alas membranosas que en reposo se pliegan sobre el
abdomen. Los adultos también poseen cercos y antenas largas. En las
unidades triásicas argentinas que registran faunas de insectos
(formaciones Potrerillos y Los Rastros) sólo se han encontrado
insectos terrestres o formas adultas terrestres. El material fue
coleccionado en 1979, durante los trabajos de
tesis en la ladera sur del Cerro Cacheuta, Mendoza
(Argentina). Este espécimen representa el primer registro de un
estado inmaduro acuático en la entomofauna triásica de la Argentina.
El espécimen presenta un muy buen estado de preservación ya que esta
prácticamente completo. En la cabeza, se observan detalles como las
mandíbulas, las antenas, un palpo, los ojos y la sutura fronto-clipeal.
En el tórax, se observa la zona esternal, las coxas, el fémur de los
tres pares de apéndices, y solo en algunos de ellos se puede ver la
región basal de las tibias.
Notopamphagopsis
bolivari.
Cabrera 1928.
|
 |
|
 |
|
Ala de
Notopamphagopsis
bolivari (entre la marca blanca) recuperada
en el Cerro Cacheuta, Provincia de Mendoza, Argentina.
Exhibida en el Museo de La Plata. |
|
Posible aspecto de
Notopamphagopsis
bolivari (*). |
Insecto,
Orthoptero.
Los insectos se fosilizan rara vez debido a que carecen de esqueleto
duro. Usualmente solo se conservan las alas, siempre que existan las
condiciones favorables como los que ofrecen los barros finos de los
lagos y pantanos.
Notopamphagopsis
bolivari es un Orthoptero emparentado con los saltamontes
actuales. Este antecesor paleozoico superior no ha sufrido
modificaciones morfológicas substanciales en los últimos 250
millones de ańos. Tienen un
aparato bucal masticador. La cabeza es por lo general grande y
redonda, a veces cónica (Acrida), bastante móvil en el
protórax, hipognata, con un robusto aparato masticador; los palpos
maxilares están constituidos por cinco segmentos, los labiales por
tres. Los ojos compuestos son hemisféricos o hemiovoidales. A menudo
están presentes tres ocelos. Las antenas, insertadas entre los ojos,
son filiformes, rara vez pectinadas. Existen en diferentes
proporciones. El protórax es siempre más grande que los demás
segmentos, y presenta un amplio pronoto que se extiende hacia atrás,
a veces excediendo la extremidad del abdomen y se dobla lateralmente
hasta cubrir por completo las pleuras protorácicas. El mesotórax, el
metatórax y el primer segmento del abdomen se encuentran soldados
entre sí. Las alas anteriores (Tegminas) son estrechas, alargadas,
del mismo color que el resto del cuerpo y con las nervaduras
onduladas. Los restos
fósiles de
Notopamphagopsis bolivari
proceden de la Formación Los Rastros (cuenca del Bermejo,
La Rioja, Argentina) y de la Formación Potrerillos en la Quebrada
del Durazno (Sur del Cerro Cacheuta), Provincia de Mendoza,
Argentina. Otros insectos conocidos de esta área;
Argentinosyne duraznoensis y
Cacheutacicada kurtzae.
Myriolepis elongatus.
Cabrera 1944.
| |
|
 |
 |
|
Ejemplar original de M.
elongatus. Este ejemplar fue utilizado para identificar la
especie. Museo de La Plata. |
Posible aspecto de los
peces condrosteos. |
Pez. Se trata de un
representante del primitivo grupo de peces llamado Chondrosteos,
caracterizados principalmente por un fuerte recubrimiento de escamas
romboidales, con la cola tipo primitivo, heterocerca, o sea, formada
por un lóbulo principal de ubicación dorsal en las formas
primitivas, con otros rasgos primitivos en las aletas, cintura
pectoral y cráneo. Los peces condrosteos se los ha registrado en
distintos continentes, tanto en capas de depósitos marinos como
continentales, desde el Devonico en Adelante. Durante el Triasico la
protagonizaron una gran diversificación, dando lugar al surgimiento
de no menos de 7 ordenes nuevos, que en su evolución se aproximaron
automáticamente a los tipos morfológicos de otros peces mas
avanzados como los Neopterigios. El material procede de la formación Los
Rastros, en Ischigualasto. La pieza tipo se encuentra parcialmente
preservado, con el cráneo y la región caudal con algunas faltantes.
No obstante presenta buenos detalles de su recubrimiento de escamas
y alguna información sobre la posición de las aletas dorsales,
pectorales y pélvicas. El tamańo de la parte conservada es de unos
45 centímetros, y completo el ejemplar debió medir unos 60
centímetros de largo total. Su dieta era variada, desde pequeńos
organismos y peces de tamańo menor este. También fueron encontrados
en el Triasico de Mendoza y en el triasico de Australia (Myriolepis
latus).
Challaida magna.
Rusconi 1949.
|
 |
|
 |
|
Ejemplar de Challaia
magna, en el Museo de Ciencias Naturales y
Antropológicas Juan Cornelio Moyano de la
ciudad de Mendoza. |
|
Posible aspecto de Challaia
magna (*). |
Pez. Actinopteri
del orden palaeonisciformes, del triasico superior de la formación
Cacheuta en la provincia de Mendoza. Todos ellos tienen escamas
ganoideas de forma romboidal, una única aleta dorsal y una aleta
caudal heterocerca. La característica principal de los
actinopterigios es la posesión de un esqueleto de espinas óseas en
sus aletas. De hecho, el término Actinopterygii significa "aletas
radiadas". Tienen el cráneo cartilaginoso (en parte calcificado)
recubierto por huesos dérmicos, y un solo par de aberturas
branquiales cubiertas por un opérculo. También se caracterizan por
presentar escamas ganoideas (carácter autapomórfico, es decir,
exclusivo del grupo) que presentan ganoína, tejido óseo esponjoso y
tejido óseo laminar. En los peces actuales la escama ganoidea se
reduce (leptoidea) presentándose dos tipos: cicloideas y ctenoideas,
en las cuales sólo se presentan tejido óseo laminar, sin ganoína ni
esponjoso.
Neochallaia tellecheai.
Rusconi 1948.
|
 |
|
 |
|
Ejemplar de
Neochallaia tellechei del Cerro
Las Cabras,
precordillera de la provincia de Mendoza.
Museo de Mendoza. |
|
Posible
aspecto de Neochallaia tellechei (*). |
Pez.
Vivió durante el
triasico medio, y hallado en sedimentos de la formación Cerro Las
Cabras, en la zona de la precordillera de la provincia de Mendoza.
Fue recuperado por Rusconi y posteriormente estudiado por Lopez
Arbarello entre otros.
Eran peces
condrósteos (Chondrostei) es una de las dos subclases de la clase
Actinopterygii de peces óseos. Proceden de peces primitivos
cartilaginosos que muestran osificación. Por ello, su nombre viene
del griego chondros, que significa. A este grupo hace tiempo
se les clasificaba dentro de los tiburones: las similitudes eran
obvias, no sólo la pérdida de hueso en la mayoría de los condrósteos,
también la estructura de la mandíbula es más parecida a la de los
tiburones que a la de otros peces óseos, y en ambos faltan las
escamas (excluyendo a los esturiones). Sin embargo el registro fósil
sugiere que estos peces tienen más en común con los teleósteos de lo
que apariencia externa parece sugerir. Características comunes
adicionales incluyen los espiráculos (en esturiones) y la cola
heterocerca (la extensión de las vértebras por el interior del
lóbulo más grande de la aleta caudal.
Calaichthys tehul.
Gouiric Cavalli, S. 2017.
|
 |
 |
 |
|
Craneo y aleta de Calaichthys tehul. Prensa. |
Detalle de las
escamas de Calaichthys tehul. Prensa. |
Aspecto en
vida del pez del Triasico de Mendoza. Prensa. |
Pez.
Los restos de un pez de 240 millones de ańos, que vivió en el
Triásico medio y perteneció a una especie de la que no se tenía
registro en Sudamérica, fueron descubiertos en la localidad de
Potrerillo por un equipo de investigadores del Conicet. Se trata de un
pez del grupo de los "Redfieldiiformes" que habitó el planeta en tiempos
de Gondwana, el bloque continental meridional desprendido del
supercontinente Pangea que al escindirse dio origen, entre otras cosas,
a la Antártida y Sudamérica. En ese entonces, lo
que hoy se conoce como "Mendoza" se cubría de lagos efímeros, es decir,
que se secaban rápidamente por la calidez del clima, y luego de la mayor
extinción de especies que se registró en el planeta, esos espacios
acuáticos fueron poblados por una gran diversidad faunística. La campańa de los
expertos del Instituto Argentino de Nivología, GlacioIogía y Ciencias
Ambientales (Ianigla, Conicet - UNCU - Mendoza) y el Museo Argentino de
Ciencias Naturales "Bernardino Rivadavia" tenía como objetivo la
recolección de plantas, "palinomorfos" e insectos de ese tiempo
histórico. Pero durante las excavaciones se encontraron con lajas
que contenían pequeńos peces. Debido a su larga historia evolutiva
muchos órdenes que aparecen representados en el registro fósil y que
hoy están completamente extintos no tienen una relación de
parentesco con las formas actuales. Es el caso de este nuevo
ejemplar, que tiene características compartidas con peces
paleozoicos y con otros más modernos. En los ejemplares se
distinguen las aletas pectorales, pélvicas, dorsal y anal, en las
que aparecen estructuras similares a las escamas que están en su
borde anterior. "Habrían servido para favorecer la hidrodinamia. Son características en las aletas pares
e impares de muchos peces paleozoicos y mesozoicos, pero no están
presentes en los de la actualidad", apuntó. Casi todos los ejemplares
que la experta estudió en el museo local tienen intacta la cabeza, en la
que se pueden distinguir fácilmente los grandes ojos, la mandíbula
superior e inferior y el opérculo, que es el límite entre el cráneo y el
tronco.
Botánica
del Triasico.
|

Cladophlebis,
una planta muy frecuente en el registro fosilífero de
Ischigualasto. |
Fronde fértil
de Asterotheca de la formación Los Rastros. |

Impresiones de
frondes fértiles de Asteroyheca s.p.
|
Paleobotanica.
El Triasico del Valle
de la Luna en la Provincia de San Juan, Republica Argentina, es uno
de los pocos sitios del mundo donde se han hallado restos botánicos
bien preservados pertenecientes al triasico. A continuación te
mostraremos algunas fotografías de estos fósiles, pero si deseas
mas información, te aconsejamos comprar el libro "El Triasico
de San Juan y La Rioja, Argentina y sus Dinosaurios" del Doctor
José Bonaparte (1997) de la Editorial del Museo Argentino de
Ciencias Naturales de Buenos Aires. No existían en ese entonces las
plantas con flores (Angiospermas) y los vegetales dominantes eran un
grupo extinguido de Gimnospermas primitivas llamadas Coristospermas,
algunas de porte arbórea y muy abundantes como Dicroidium.
Dicroidium
odontopteroides.
Morris in
Strzelecki 1845.
|
 |
 |
 |
|
Dicroidium
(Carystopermaceae) un árbol Triasico de Argentina. |
Otra especie de
flora de Dicroidium de Los Rastros. |
Característica de la flora del
Triasico de Ischigualasto. |
Paleobotanica.
Las comunidades vegetales del Triasico de
Patagonia cambian drásticamente con respecto a aquellas del
Paleozoico superior. Esta nueva asociación florística domina esta
región al igual que en Ischigualasto. La planta mas importante es
una con forma de helecho que lleva semillas y se denomina Dicroidium.
La misma pertenece a un grupo de angiospermas sumamente primitivas
que combina una serie de características como la presencia de óvulos
con hojas sumamente divididas. Hay importantes asociaciones con
otras especies de helechos primitivos y coniferas, en fin, toda una
gama rica y exuberante la cual muy probablemente constituyeron el
principal alimento de los grandes vertebrados de este periodo.
También se han encontrado una gran variedad de Dicroidium
odontopteroides en el Triasico de la pre-cordillera en
Cacheuta, Mendoza.
Sphenobaiera argentina.
Frenguelli, 1946.
|
 |
|
 |
|
Plantas fósiles del genero Sphenobaiera.
Museo de La Plata. |
|
Frondes vegetales del
Triasico superior de Cacheuta.
Museo de
Ciencias Naturales y
Antropológicas Juan Cornelio Moyano de Mendoza. |
Paleobotanica. Es un
género extinto de plantas que vivieron durante el
Triásico Superior al
Cretácico. El género Sphenobaiera se utiliza para plantas
con hojas en forma de cuńa y se puede diferenciar de
Ginkgo,
Ginkgoites y
Baiera la falta de un
peciolo.
Se extinguieron hace unos 72.600 ańos. La
familia que pertenece a este
género no es concluyente y afinidad con
Karkeniaceae había razones
morfológicas sugeridas.
Rhexoxylon brunoi.
Artabe, Brea et Zamuner 1999.
|
 |
|
 |
|
Los troncos fósiles de la formación los
colorados, en Villavicencio- Uspallata en el
Museo de
Ciencias Naturales y
Antropológicas Juan Cornelio Moyano de Mendoza. |
|
Restos de troncos en sedimentitas triásicas del área de
Paramillos de Uspallata (Cuenca Cuyana, Mendoza). Imagen del
Inst. de Estudios Andinos. |
Paleobotanica. Es un género de
coníferas de la familia Araucariaceae. Los troncos fósiles de este
genero procede de la formación los colorados, en Villavicencio-
Uspallata, en Mendoza. Son principalmente árboles grandes con un
vástago erguido masivo, alcanzando una altura de 30 a 80 m. Las
ramas suelen ser horizontales, y bien separadas; estando cubiertas
con hojas coriáceas o aciculares. Charles Robert Darwin (1809/1882)
recorrió la provincia de Mendoza (Argentina) durante 1835, como
parte de aquella histórica travesía alrededor del mundo (1831/1836)
a bordo del Beagle, expedición dirigida por Fitz Roy. Entre sus
descubrimientos más notables, y a la vez poco recordados, se
encuentra un bosque de árboles petrificados de más de sesenta
millones de ańos de antigüedad. El autor de “El Origen de las
Especies” describe en sus diarios de viaje las araucarias
fosilizadas encontradas en el desértico y serrano paraje de Los
Paramillos, próximo a la localidad de Uspallata, hoy Departamento de
Las Heras. Darwin describió el perfil de la Sierra de Uspallata y,
con su agudo poder de observación, describió una secuencia
sedimentaria con más de 52 troncos petrificados en posición de vida.
Los describió en detalle, al igual que los sedimentos circundantes,
caracterizados por su alto contenido volcánico.
Dictyophillum tenuifolium.
Stipanicic y Menéndez, 1949.
 |
|
 |
|
Dictyophillum tenuifolium, del Triásico Superior del
Paso Flores (Provincia de Río Negro). Paseo Paleontológico
de Bariloche. |
|
Aspecto de un representante de la Familia Dipteridaceae.
|
Paleobotanica. Los
helechos de la Familia Dipteridaceae constituyen uno de los grupos
sistemáticos más reconocibles en las floras mesozoicas debido a su
característica venación anastomosada. Aunque este carácter es
compartido por otros helechos (e.g., Matoniaceae y géneros
como Weichselia del Mesozoico y Polypodiaceae del
Cenozoico), de todas maneras son los más típicos y ampliamente
difundidos en el Mesozoico del mundo.
Dictyophillum tenuifolium, es una especie,
originalmente descrita por Stipanicic y Menéndez (1949) del Triásico
Medio de la Formación Barreal de la Provincia de San Juan
(Argentina), fue luego enmendada por Bonetti y Herbst (1964) con
materiales fértiles del Triásico Superior del Paso Flores (Provincia
del Neuquén, Argentina). Es una especie muy característica en cuanto
a su morfología. En el estudio de las Dipteridaceae del Lías de
Piedra Pintada (Neuquén, Argentina), Frenguelli (1941) seńaló, en
los comentarios de Dictyophyllum rothi Frenguelli, la
notable afinidad morfológica que tiene lugar en varias especies de
Europa y Oriente y admitió que es muy difícil llegar a definir
especies cuando sólo se dispone de fragmentos de hojas. Como
consecuencia, Frenguelli (1941) también expresó su escepticismo
respecto a la identificación taxonómica de los vegetales triásicos
de La Ternera (Chile), indicada por Zeiller (1875) y Solms-Laubach
(1899). El registro fósil comienza en el Triásico tardío.
Actualmente distribuidas en India, el sudeste de Asia, el este y
sudeste de China, centro y sur de Japón, y desde Malasia hasta
Melanesia y el oeste de Polinesia (Samoa).
Zuberia feistmanteli.
(Johnston) Frenguelli emend.
Artabe 1990.
|
 |
|
 |
|
Frondes vegetales de
Corystospermales del Triasico de Barreal, San Juan. Exhibido en el
Museo de Cs Nat. y Antropológicas de Mendoza. |
|
Aspecto de un
bosque triasico de
Corystospermales (*): |
Paleobotanica. Este
genero y especie con abundante presencia en Cacheuta
(Luján de Cuyo)
en la provincia de
Mendoza, como así también en San Juan, corresponde a las conocidas
plantas Corystospermales, un orden de plantas que aparecieron en el
Pérmico y se extinguieron en el Cretácico. Se trata de un grupo
relativamente pequeńo de plantas con distribución principalmente en
Gondwana. Fueron las plantas dominantes del hemisferio sur durante
el Triásico, y se utilizan en la bioestratigrafía y paleoecología de
ese periodo. Se caracterizan por tener un raquis principal bifurcado
en sus frondes. Los tallos presentaban verdadero crecimiento
secundario en "cuńas" de xilema centrífugo y centrípeto. Los órganos
de reproducción femeninos son tipo cúpula, que surgen de
braquiblastos (pequeńas ramas abortadas). Los órganos masculinos son
sinngios y presentan un polen disacado. Otros géneros conocidos en
el Triasico de Cacheuta; Johnstonia, Xylopteris, Scytophyllum,
Pachydermophyllum, Johnstonia, Dicroidium, Cladophlebis, Asterotheca,
Nododendron, Neocalamites, Equisetites entre otros.
Baiera furcate.
Lindley and Hutton, 1837.
|
.jpg) |
|
.JPG) |
|
Improntas de
Baiera furcate. (*): |
|
Baiera
furcate, procedente de Los Potrerillo, Mendoza y
exhibida en el Museo de La Plata. |
Paleobotanica. Es
un género de gimnospermas prehistóricas en el orden Ginkgoales. Está
relacionado con los géneros Ginkgo y Ginkgoites. Se lobula en cuatro
segmentos y no tiene tallo. Los fósiles de Baiera han
sido conocidos desde el Pérmico hasta el Cretácico. Las Ginkgoaceae —una familia de Ginkgoales—
son gimnospermas arbóreas con un único representante vivo actual,
del género Ginkgo y especie Ginkgo
biloba. A diferencia de las cícadas, y como casi todas
las demás gimnospermas, son árboles leńosos altamente ramificados.
Puede ser reconocido por el hecho de que tiene ramas cortas además
de ramas largas. De hasta 30 metros de altura, con una corona más o
menos asimétrica y corteza gris y surcada. Sin canales de resina.
Hojas simples, de disposición espiralada, y ampliamente espaciadas
en largas ramas cerca de las puntas de las ramas, también
ampliamente empaquetadas en ramas cortas en los árboles más viejos,
con forma de abanico, bilobadas o enteras, deciduas y amarillas
brillantes en el otońo, de venación dicotómica. En Argentina se han
recuperado fósiles en la Formación Paso Flores, en la localidad
Cańadón de Pancho, área occidental del río Collón Curá, Provincia
del Neuquén, Cacheuta y Los Poterillos en la pre-cordillera en la
Provincia de Mendoza Otras especies conocidas son; Baiera cuyana
y Baiera gracilis.
Nododendron
suberosum. Artabe y Zamuner
1991.
|
Ejemplares de
Nododendron suberosum. Proceden de la formación
Potrerillos, Triasico de la Provincia de Mendoza. Exhibidos
en el Museo de La Plata |
Paleobotanica. Este
taxón fue descripto e ilustrado para la Formación Potrerillos
expuesta en el cerro Cacheuta. En la Formación Paramillo, localidad
de Agua de la Zorra provincia de Mendoza, se hallaron restos afines
a Nododendron
suberosum en asociación estrecha con
Neocalamites carrerei y estructuras reproductivas (Neocalamostachys
arrondoi). La familia equisetáceas es monotípica (con un
solo género), siendo su género viviente, Equisetum
(conocido como cola de caballo), que es el único sobreviviente hasta
la actualidad de todas las equisetópsidas, las cuales tienen un
extenso registro fósil entre el Devónico y el Triasico. Los
equisetos son plantas vasculares
con ciclo de vida
haplodiplonte donde la alternancia de generaciones es bien
manifiesta, con esporofito y gametofito multicelulares e
independientes, con esporas como unidad de dispersión y de
resistencia. Los tallos, fotosintéticos, crecen a partir de un
rizoma subterráneo mediante un meristema que consta de una célula
apical piramidal que se divide en tres direcciones.
Concostrácos.
|
 |
|
 |
|
Concostrácos encontrados en el Cerro de
Cacheuta, provincia de Mendoza y exhibidos en el Museo de La
Plata. |
|
Aspecto y forma de vida de los
Concostrácos.
Museo de Ciencias Naturales de la Univ. Nacional de Salta. |
Invertebrados. Los concostráceos se
caracterizan por tener el cuerpo protegido por un caparazón bivalvo
(dos conchas unidas por una membrana que permite cierta
articulación). Este escudo les hace asemejarse mucho a las almejas,
con las cuales comparten la característica de líneas concéntricas en
cada concha. La articulación de las conchas permite, que cuando los
músculos están relajados, emerjan las patas y las antenas, las
cuales usa para remover el agua y llevar el alimento a la boca. En
caso de un ataque, los músculos abductores se contraen y las valvas
se cierran. Las demás características de los concostráceos son
similares a las de los branquiópodos. La conchostracofauna
previamente conocida de la cuenca Cuyana provenía de las formaciones
Cerro de las Cabras, Potrerillos y Cacheuta, Triasico de Mendoza. En
un ambiente netamente fluvial de tipo efímero y en las áreas más
deprimidas del sistema, se desarrollaron extensos cuerpos de agua
semipermanentes, lagunas localizadas y/o pantanos producidos por el
desbordamiento de los canales o flujos en mantos, que permitió el
desarrollo de estos organismos. Actualmente se considera un grupo
parafilético y ha sido escindido en los subordenes Laevicaudata,
Spinicaudata y Cyclestherida, dentro de la clase Branchiopoda.
(*) Imagen de autor u origen desconocido.
Ver sobre nuestras
Políticas de uso de imágenes.
|
Bibliografía Utilizada.
Arcucci, A.B.
& Rodolfo A.C. 2003.
A new Triassic carnivorous dinosaur from
Argentina.
Ameghiniana
40(2):217-228.
Bonaparte, J.F (1963). "Promastodonsaurus
bellmanni n. gen. et n. sp., capitosa urido del Triasico medio
de Argentina (Stereospondyli, Capitosauroidea)". Ameghiniana 3: 67–78.
Bonaparte, J.F. (1969) Dos
nuevas faunas de reptiles triásicos de Argentina. IST
Gondwana Symp., Mar del Plata Ciencias Tirra.
Bonaparte, J.F. 1975.
Nuevos materiales de Lagosuchus talampayensis Romer (Thecodontia-Pseudosuchia)
y su significado en el origen de los Saurischia: Chańarense
inferior, Triásico medio de Argentina. Acta Geológica Lilloana
13:5-90.
Carroll, Robert
L. (1988). Vertebrate Paleontology and Evolution. W.H.
Freeman and Company.
Casamiquela, R. M. 1980.
La presencia del género Plateosaurus (Prosauropoda) en el
Triásico superior de la Formación El Tranquilo, Patagonia.
Actas II Congreso Argentino de Paleontología y
Bioestratigrafía. I Congreso Latinoamericano de
Paleontología, 1, 143-158.
Desojo, J.B.; Ezcurra, M.D. (2011).
"A reappraisal of the taxonomic status of Aetosauroides (Archosauria,
Aetosauria) specimens from the Late Triassic of South America and
their proposed synonymy with Stagonolepis". Journal of
Vertebrate Paleontology 31 (3): 596–609.
Gallego, O.F., Martins-Neto,
R.G., Rébori, L.O., Dicindio, P.A. & Sallenave, S. 2006.
Ninfa de Plecoptera (Insecta) del Triásico del Cerro
Cacheuta (Provincia de Mendoza), Argentina. Reunión de
Comunicaciones Científicas y Tecnológicas, Secretaría General
de Ciencia y Técnica, Universidad Nacional del Nordeste,
Resistencia, 24-27
José Bonaparte. El Triásico de San Juan-La
Rioja Argentina y sus dinosaurios. Buenos Aires. Museo Argentino de
Ciencias Naturales “Bernardino Rivadavia”, 1997, 190 páginas.
Lecuona, A. & J. B.
Desojo. 2011. Hind limb osteology of Gracilisuchus stipanicicorum
(Archosauria: Pseudosuchia). Earth and Environmental Science
Transactions of the Royal Society of Edinburgh, 102, 105–128.
Max C. Langer and Cesar L. Schultz (2000). «A new species of the
Late Triassic rhynchosaur Hyperodapedon from the Santa Maria
Formation of south Brazil». Palaeontology 43 (6):
633–652.
Magnussen Saffer,
Mariano (2010). Algunas
curiosidades de la fauna triásica (Therapsida, Archosauria y
Labyrinthodontia) de Ischigualasto.
Paleo, Boletín Paleontológico.
Ańo 8. 49: 11-15.
Magnussen Saffer,
Mariano (2010).
Conociendo a los Dinosaurios Triasicos de la Republica
Argentina. Una ventana a los posteriores gigantes. Paleo,
Revista Argentina de Paleontología. Boletín Paleontológico.
Ańo 8. 50: 28 -36.
Martínez,
Ricardo N. y Alcober, Oscar A. (2009).
«A basal sauropodomorph (Dinosauria:
Saurischia) from the Ischigualasto Formation (Triassic,
Carnian) and the early evolution of Sauropodomorpha». PLoS
ONE 4 (2). ISSN , 1-12.
Marsicano C. A. (1990)
Redescripción del cráneo del holotípo de Pelorocephalus
mendozensis Cabrera, 1944 (Amphibia, Temnospondyli) del Triásico
del Cerro Bayo, Provincia de Mendoza. Ameghiniana. vol. 27,
no1-2, pp. 39-44 (32 ref.)
Palmer,
D., ed. (1999). The Marshall Illustrated Encyclopedia of
Dinosaurs and Prehistoric Animals. London: Marshall Editions.
p. 193.
Ricardo N.
Martínez, Paul C. Sereno, Oscar A. Alcober, Carina E.
Colombi, Paul R. Renne, Isabel P. Montańez and Brian S.
Currie (2011).
«A Basal Dinosaur from the Dawn of the
Dinosaur Era in Southwestern Pangaea». Science 331 (6014):
206–210.
Romer, A. S. (1971). "The Chańares
(Argentina) Triassic reptile fauna. XI. Two new long-snouted
thecodonts, Chanaresuchus and Gualosuchus".
Breviora 379: 1–22.
Rougier, G. W., Fuenta de la
M. S., and Arcucci A. B., 1995, Late Triassic Turtles from
South America: Science, v. 268, p. 855-858.
Sebastián Apesteguia. Vida en Evolución, la historia natural
vista desde Sudamérica”.- 2010.
Sereno, Forster, Rogers and Monetta, 1993.
Primitive dinosaur skeleton from Argentina and the early
evolution of Dinosauria.
Nature. 361,
64-66.
Ver mas
bibliografía utilizada para hacer la presente Pagina;
AQUÍ.
Ver bibliografía de divulgación sugerida;
AQUÍ. |
|